

Незначне старіння — темп старіння, який важко статистично відрізнити від нуля у масштабах даної вибірки, а також «нестаріння» — нульову кореляцію між віком й імовірністю смерті. Іншими словами, мова йде від випадків потенційного безсмертя до видів істоти яких демонструють величезну максимальну тривалість життя (МТЖ), через що ми не можемо візуально спостерігати ознак їх старіння. Цей термін вперше введено Калебом Фінчем у 1990 році.
Сутність процесу
У 2001 році Фінч і Остед запропонували мінімальні критерії для віднесення того чи іншого біологічного виду до категорії видів для яких характерне незначне старіння: відсутність вікового зростання темпу смертності і захворюваності після статевого дозрівання, зниження темпу розмноження і ряд фізіологічних показників. Такі істоти повинні старіти так поволі, що зафіксувати будь-які вікові зміни було б практично неможливо. На сайті AnAge є перелік видів для яких характерне незначне старіння.
У цьому переліку є морський окунь алеутський (Sebastes aleutianus);— МТЖ 205 років; черепахи: Chrysemys picta;— МТЖ 61 років; Emydoidea blandingii;— МТЖ 77 років; Terrapene carolina ;— МТЖ 138 років; морський їжак Strongylocentrotus franciscanus ;— МТЖ 200 років; двостулковий молюск Arctica islandica ;— МТЖ 400 років; сосна остиста (Pinus longaeva) — МТЖ 4731 років.
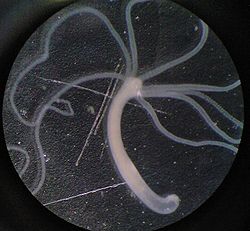
Безумовно, список бази даних AnAge неповний, і його необхідно поповнити гідрою Cnidaria vulgaris, так як потенційне безсмертя цього виду кишковопорожнинних доведене Даніелем Мартінезом у 1998 році. Також потенційно безсмертна медуза Turritopsis nutricula . Гідроїдні — модульні організми на стадії поліпу, але медузоїдна стадія унітарна. Більшість медуз після завершення репродуктивного циклу гинуть, але Turritopsis nutricula повертається до ювенальної стадії — модулярного поліпу, уникаючи смерті. Такий цикл Turritopsis nutricula може повторювати нескінченно, що робить її потенційно безсмертною. Окрім того, ряд сторінок згаданої вище бази даних AnAge присвячені видам губок, які демонструють рекордне довголіття не тільки серед тварин, але і серед інших живих істот. Наприклад, рекорд тривалості життя серед Animalia демонструє екземпляр антарктичної губки Anoxycalyx joubini, вік якої оцінюють від 15 до 23 тис. років.
У науковій літературі часто згадують види щук, осетерів, білугу (Husa husa), а також групер — кораловий лосось (Plectropomus pessuliferus), гігантський групер (Epinephelus lanceolatus) американський омар (Homarus americanus) і т. д. як такі для яких характерне незначне старіння. Можливо видів із незначним старінням набагато більше ніж тих, про які відомо сучасній біологічній науці.
Недавно факт незначного старіння серед прісноводих двостулкових молюсків встановив російський дослідник В. В. Зюганов:— прісноводна річкова скойка (Margaritifera margaritifera) має найдовшу тривалість життя серед прісноводих безхребетних тварин — МТЖ 210 — 250 років і для неї характерне незначне старіння.
Більш того, і серед наземних рослин (Embryophyta) значна частина видів демонструє незначне старіння. Рекорд тривалості життя належить екземпляру Posidonia oceanica— його вік оцінюють в 100 000 років . Друге місце належить клонам, які протягом тисячоліть не втрачають своїх ювенальних якостей, наприклад, клон Lomatia tasmanica має вік 43 тис. років. Деякі дерева — баобаби, драконові дерева, ліванські кедри і т. д. живуть тисячі років, тому частину деревних форм відносять до категорії видів для яких характерне незначне старіння.
Сучасні погляди на природу та механізми незначного старіння
Перераховані у списку чинники провокують старіння, скорочуючи життя всім живим істотам, окрім тих видів для яких характерне незначне старіння.
- мутації генів
- оксиданти
- укорочення теломер
- метилювання ДНК
- глікозування білків і ДНК
- нестабільність генома
- гормональний дисбаланс
- канцерогенез
- шкідливий вплив стресорних факторів.
Чому організм видів для яких характерне незначне старіння нечутливий до таких небезпечних загроз — думка сучасних дослідників розходиться.
В. В. Зюганов вважає, що як правило, види тварин із незначним старінням мають безперервний асимптотичний ріст і їх репродуктивна здатність зазвичай не зменшується, а навпаки, збільшується з віком, оскільки позитивно корелює з розмірами тіла. Узагальнення польових спостережень приводить до висновку, що тварини із незначним старінням не вмирають від випадкових причин, як вважає ряд геронтологів, а гинуть з голоду або по-суті від геометричних наслідків свого безперервного росту — надкрупних розмірів, що призводить до виходу з своєї екологічної ніші — втрати необхідної спритності у добуванні їжі і як фінал — від дефіциту поживних речовин. Як доказ В. В. Зюганов дає дуже цікавий приклад з гігантським групером (Epinephelus lanceolatus), який має МТЖ близько 100—120 років.
|
Спостереження в Індійському океані за харчовою поведінкою цих велетнів завдовжки 2.5 метра і вагою до 400 кг показали, що гігант програє конкуренцію із молодими особинами в охороні індивідуальних ділянок. Якщо поряд молодий самець, то він атакує старенького, тому що спритніший . Старий підтискає хвіст і тікає. А старенькому так хотілося скуштувати краба, але на своїй території він це вже зробити не може. Результат - програш унаслідок конкуренції. Тобто, стареньким або їжі не вистачає, або накопичуються негативні ефекти від стресів. Старенькому не вистачає проворності дати здачі молодому конкурентові через свої дуже крупні розміри, а також через крупні розміри він не може залізти у печерку відпочити. Фактично, він виростає зі своєї екологічної ніші, але не старіє. У нього прекрасний зір, він бачить найменше ворушіння вусиків креветки з-під будь-якого каменя. |
А. В. Макрушин, один із співавторів гіпотези еволюційного виникнення процесів старіння та онкогенеза вважає, що первинний механізм старіння виник на «колоніально-сидячому» етапі еволюції Metazoa. Модульний організм — донорно-акцепторна система у якої процеси ембріогенеза і старіння відбуваються по життєво і одночасно. У сидячих колоніальних видів Metazoa руйнування важливих органів — складова частина нормального онтогенезу, що супроводжує ітеропарне і семельпарне безстатеве розмноження. Старий модуль може відмерти, віддаючи ресурси новому. Поживні речовини від одного модуля передаються іншим, що може детермінувати локальне старіння «непотрібного» модуля через потребу колонії міняти форму через зміну оточення. Цей процес супроводжує інволюція паренхимних клітин, і зрештою смерть модуля. У модульних видів істотою, що володіє унікальним генотипом, є колонія, яка виникла із зіготи. Тому смерть модулів не прискорює еволюцію, так як призводить цю істоту до смерті, а навпаки омолоджує її. Кожен модуль, перш ніж померти через старість, породжує декілька дочірніх модулів. А. В. Макрушин робить висновок, що саме завдяки заміні старих модулів молодими, істоти перших на Землі Metazoa, так і багатьох нині існуючих модульних видів були потенційно безсмертними. А. В. Макрушин визнає факти незначного старіння у унітарних видів, але заперечує можливість у них випадків потенційного безсмертя.
У наш час виникла інтрига навколо астроцитарної гіпотези старіння ссавців. Згідно з постулатами цієї гіпотези серед філогенетичних гілок хребетних, види із незначним старінням можуть бути виявлені у риб, хвостатих амфібій, черепах, і можливо, у птахів на фоні того, що заперечується можливість існування видів із незначним старінням у ссавців. Окрім того, автор астроцитарної гіпотези старіння ссавців]] О. Г. Бойко також не визнає можливість існування видів із незначним старінням і серед ряду філ безхребетних: Appendicularia, Nematoda, Rotifera і Insecta так як для них характерна елімінація стовбурових клітин у організмі дорослих особин після закінчення ембріогенезу . У 2008 році група Рошель Баффенштейн повідомила про факт незначного старіння серед ссавців — кротячого пацюка (Heterocephalus glaber) . Але цей факт був поставлений під сумнів Калебом Фінчем, як такий, що не відповідає критеріям незначного старіння. Також цей факт не визнав автор астроцитарної гіпотези старіння ссавців. З російських учених за групу Рошель Баффенштейн «хворіють» академік В. П. Скулачов та наймолодший доктор наук Росії Олексій Москальов, який також є опонентом астроцитарної гіпотези старіння ссавців.
У багатьох країнах світу створені центри для дослідження феномена незначного старіння.
Таким чином, старіння — не обов'язковий атрибут існування багатоклітинних організмів, оскільки багато видів можуть чудово існувати без нього . Смерть від старості є винахід еволюції, і саме у цьому велика правда Августа Вейсмана.
Література
- Finch C.E., 1990. Longevity, Senescence, and the Genome. Chicago: Univ. of Chicago Press. 922 p.
- Finch C.E., Austad S.N., 2001. History and prospects: symposium on organisms with slow aging // Exp. Gerontol. V. 36. № 4-6. P. 593—597.
- Human Aging Genomic Resources. Aging, longevity, and life history of Scolymastra joubini. 2007.
- Зюганов В. В., 2004. Арктические долгоживущие и южные короткоживущие моллюски жемчужницы как модель для изучения основ долголетия // Успехи геронтол. Т. 14. С. 21-31
- Зюганов В.В. (2008). НЕСТАРЕЮЩИЕ ЖИВОТНЫЕ. ПОЧЕМУ ОНИ ЖИВУТ ДОЛГО, НО НЕ ВЕЧНО?. Использование и охрана природных ресурсов в России (информационно-аналитический бюллетень) № 2 (98). С. 30-36. Архів оригіналу за 4 вересня 2011. Процитовано 11 грудня 2010.
- Макрушин А. В. Как и почему возникли механизмы старения и онкогенеза: гипотеза // Журн. общ. биол. 2008. Т. 69. № 1. С. 19-24.
- Макрушин А. В. Первичный механизм старения: гипотеза // Успехи геронтол. 2006. Т. 19. С. 25-27.
- Макрушин А. В. Эволюционные предшественники онкогенеза и старческой инволюции // Успехи геронтол. 2004. Т. 13. С. 32-43.
- Макрушин А. В. Как мог возникнуть механизм старческой инволюции // Успехи геронтол. 2001. Т. 7. С. 50-51.
- Макрушин А. В., Худолей В. В. Опухоль как атавистическая адаптивная реакция на условия окружающей среды // Журн. общ. биол. 1991. Т. 52. № 5. С. 717—722.
- Buffenstein R. Negligible senescence in the longest living rodent, the naked mole-rat: insights from a successfully aging species // J. Comp. Physiol. B. 2008. Vol. 178. P. 439—445.
- Finch C.E. 2009. Update on Slow Aging and Negligible Senescence — A Mini-Review // Gerontology 55:307-313
- Boyko O.G. (2004). Do mammals die young!? An age-dependent mechanism of mammals self-destruction. Ukr. Bioorg. Acta 1–2: 3–12.
- Бойко А.Г. (2007). Дифференцировка клеток радиальной глии в астроциты - вероятный механизм старения млекопитающих. Журнал общей биологии. 68. № 1.: 35–51.
- Бойко А.Г., Лабас Ю.А., Гордеева А.В. (2010). Очерк филогенетической истории феномена старения Metazoa (К вопросу создания общей теории старения metazoa). Успехи геронтол. 23. № 1.: 21 – 29.
- Москалев А.А. (2010). Эволюционные представления о природе старения Дифференцировка клеток радиальной глии в астроциты - вероятный механизм старения млекопитающих. Успехи геронтол. 23. № 1.: 9 – 20.
- Guerin, J. 2004. Emerging area of aging research: long-lived animals with «negligible senescence» // Ann N Y Acad Sci. 1019:518-520.
Джерела
| |||||||||||