
Філогенетична номенклатура — це метод номенклатури таксонів у біології, який використовує філогенетичні визначення для назв таксонів, як пояснюється нижче. Це відрізняється від традиційного підходу, згідно якому назви таксонів визначаються типом, який може бути зразком або таксоном нижчого рангу, і описом у словах. Філогенетична номенклатура в даний час регулюється Міжнародним кодексом філогенетичної номенклатури (PhyloCode).
Визначення
Філогенетична номенклатура пов'язує назви з кладами, групами, що складаються з предка та всіх його нащадків. Ці групи можна еквівалентно назвати монофілетичними . Існують різні способи визначення предка, які обговорюються нижче. Після визначення предка створюють назву таксону: предок і всі організми, які є його нащадками, включаються до названого таксону. Для перерахування всіх цих організмів (тобто надання повного опису) потрібно знати повне філогенетичне дерево. Практично існує лише одна або декілька гіпотез щодо правильного дерева. Різні гіпотези призводять до того, що різні організми вважаються включеними до названого таксону, але не впливають на те, до яких організмів ця назва насправді відноситься. У цьому сенсі назва є незалежною від перегляду теорії.
Філогенетичні визначення назв клад
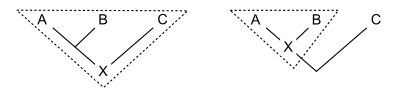
Філогенетична номенклатура пов'язує назви з кладами, групами, що складаються виключно з предка та всіх його нащадків. Тому для визначення клади потрібно лише вказати предка. Існує декілька способів як це зробити. Зазвичай на предка вказує його відношення до двох або більше специфікаторів (видів, зразків або ознак), які згадуються явно. На діаграмі показано три способи, за допомогою яких назву клади X можна визначити за допомогою вже визначених клад A, B і C:
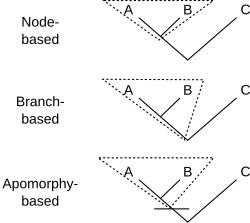
- Визначення на основі вузла може бути таким: « найближчий спільний предок A і B і всі нащадки цього предка». Таким чином, уся лінія під стиком A і B не належить до клади, до якої відноситься назва з цим визначенням.
- Приклад: Динозаври- завроподи складаються з останнього спільного предка вулканодона (A) і апатозавра (B) і всіх нащадків цього предка. Цей предок був першим завроподом. C може включати інших динозаврів, таких як стегозавр .
- Визначення на основі гілки, яке часто називають визначенням на основі стебла, читається як: «перший предок A, який також не є предком C, і всі нащадки цього предка». Таким чином, уся лінія під стиком A і B (окрім найнижчої точки) справді належить до клади, до якої відноситься назва з цим визначенням.
- Приклад: Гризуни складаються з першого предка хатньої миші (A), який також не є предком східного кролика (C), разом із усіма нащадками цього предка. Тут предком є найперший гризун. (B) є іншим нащадком, можливо, червоною білкою .
- Визначення, засноване на апоморфії, могло б звучати так: «перший предок А, який має ознаку М, успадковану А, і всі нащадки цього предка». На діаграмі M розвивається на перетині горизонтальної лінії з деревом. Таким чином, клад, до якого відноситься назва з цим визначенням, містить ту частину лінії нижче останнього спільного предка A і B, яка відповідає предкам, що мають апоморфію M. Нижня частина лінії виключається. Не обов'язково, щоб B мав ознаку M; воно могло зникнути в родоводі, що веде до B.
- Приклад: Чотириногі складаються з першого предка людини (A), від якого люди успадкували кінцівки з пальцями рук або ніг (M), і всіх нащадків цього предка. До цих нащадків належать змії (B), які не мають кінцівок.
Кілька інших альтернатив надано у PhyloCode (див. нижче).
Філогенетична номенклатура дозволяє використовувати не лише зв'язки предків, але й властивості існуючих таксонів . Один із багатьох способів специфікації класу Neornithes (сучасних птахів), наприклад, такий:
- Клас Neornithes складається з останнього спільного предка сучасних членів найбільш інклюзивної клади, яка містить Какаду жовточубого, але не динозавра Stegosaurus armatus, а також усіх нащадків цього предка.
Neornithes — це кронова група, клада, для якої останній спільний предок її існуючих членів також є останнім спільним предком усіх її членів.
Імена вузлів
- Кроновий вузол : останній спільний предок відібраного виду клади, яка представляє інтерес
- Стебловий вузол : останній спільний предок клади, що представляє інтерес , і її сестринської клади
Визначення назв парафілетичних і поліфілетичних таксонів за походженням
У PhyloCode лише клада може отримати «філогенетичне визначення», і це обмеження враховано в цій статті. Однак також можна створити визначення назв інших груп, які є філогенетичними лише за родовими зв'язками, закріпленими за видами чи зразками. Наприклад, якщо припустити, що Mammalia та Aves (птахи) визначені саме так, тоді Reptilia можна визначити як «останнього спільного предка Mammalia та Aves та всіх його нащадків, крім Mammalia та Aves». Це приклад парафілетичної групи, клада мінус одна або більше підпорядкованих клад. Назви поліфілетичних груп, що характеризуються ознакою, яка конвергентно розвинулась у двох або більше підгрупах, так само можна визначити як суму кількох клад.
Ранг
Відповідно до традиційних номенклатурних кодів, таких як Міжнародний кодекс зоологічної номенклатури та Міжнародний кодекс номенклатури водоростей, грибів і рослин, таксони, які явно не пов'язані з рангом, не можуть бути офіційно названі, оскільки застосування назви до таксону базується як на типі, так і на ранзі. Вимога до рангу є головною відмінністю між традиційною та філогенетичною номенклатурами. Це має кілька наслідків: 1) обмежує кількість вкладених рівнів, на яких можна застосовувати імена; 2) спричиняє зміну закінчень імен, якщо змінюється ранг групи, навіть якщо вона має точно тих самих членів (тобто однакову об'ємність); 3) логічно суперечить тому, що всі таксони є монофілетичними.
Особливо в останні десятиліття (завдяки прогресу в філогенетиці) таксономісти назвали багато «вкладених» таксонів (тобто таксонів, які містяться всередині інших таксонів). Жодна система номенклатури не намагається назвати кожну кладу; це було б особливо складно в традиційній номенклатурі, оскільки кожному названому таксону повинен бути наданий нижчий ранг за будь-який вже названий таксон, в який він вкладений. Тому кількість назв, які можна призначити у вкладеному наборі таксонів, не може перевищувати число загальновизнаних чинів. Жак Готьє зі співав. (1988) припустив, що якщо Reptilia присвоєно традиційний ранг класу, то філогенетична класифікація повинна присвоїти Aves ранг роду. У такій класифікації всі ~12 000 відомих видів існуючих і вимерлих птахів повинні бути включені в цей рід.
Запропоновані різні рішення зі збереженням рангових номенклатурних кодів. Patterson and Rosen (1977) запропонували дев'ять нових рангів між родиною і надродиною, щоб мати можливість класифікувати кладу оселедців, а McKenna and Bell (1997) представили великий набір нових рангів, щоб впоратися з з різноманіттям ссавців; але вони не були широко прийняті. У ботаніці, вчені з групи філогенії Покритонасінних рослин, відповідальної за найбільш поширену на даний момент класифікацію Покритонасінних, обрали інший підхід. Вони зберегли традиційні ранги родини та порядку, вважаючи їх цінними для навчання та вивчення зв'язків між таксонами, але також додали назви клади без формальних рангів.
Чинні кодекси також містять правила, за якими назви повинні мати певні закінчення залежно від рангу таксонів, до яких вони належать. Якщо група має різний ранг у різних класифікаціях, її назва повинна мати інший суфікс. Ereshefsky (1997:512) навів приклад: Simpson у 1963 році та Wiley у 1981 році погодилися, що ту саму групу родів, яка включала рід Homo, слід об'єднати в один таксон. Simpson розглядав цей таксон як родину, і тому дав йому назву «Hominidae»: «Homin-» від «Homo» та «-idae» — закінчення згідно зоологічного коду родини. Wiley вважав, що воно відноситься до рангу племені, і тому дав йому назву «Hominini», де «-ini» означає закінчення племені. Плем'я Wiley Hominini було лише частиною родини, яку він назвав «Hominidae». Таким чином, відповідно до зоологічного коду, дві групи з абсолютно однаковою межею отримали різні назви (Hominini Simpson та Hominidae Wiley), а дві групи з однаковою назвою мали різні межі (Simpson's Hominidae and Wiley's Hominidae).
У філогенетичній номенклатурі ранги не впливають на написання назв таксонів (див. Жак Готьє (1994) і PhyloCode). Проте ранги не заборонені у філогенетичній номенклатуріі, а лише відокремлені від номенклатури. Тобто не впливають на використання назв; зв'язок назв з таксонами та посилання назви на вкладені таксони .
Принципи традиційної рангової номенклатури логічно несумісні з тим, що всі таксони є строго монофілетичним . Кожен організм повинен належати до роду, наприклад, тому повинен бути рід для кожного спільного предка ссавців і птахів. Щоб такий рід був монофілетичним, він мав би включати як клас Mammalia, так і клас Aves. Однак у ранговій номенклатурі класи повинні включати роди, а не навпаки.
Філософія
Конфлікт між філогенетичною та традиційною номенклатурами відображає різні погляди на метафізику та епістемологію таксонів. Для прихильників філогенетичної номенклатури таксон — це окрема особина, сутність, яка з часом набуває та втрачає атрибути. Подібно до того, як людина не стає кимось іншим, коли її властивості змінюються через дозрівання, старість або більш радикальні зміни, такі як амнезія, втрата кінцівки чи зміна статі, таксон залишається тією самою сутністю, незалежно від того, які характеристики отримані або втрачено. Враховуючи сильні метафізичні твердження щодо неспостережуваних сутностей, зроблених прихильниками філогенетичної номенклатури, критики назвали їхній підхід есенціалізмом.
Для будь-якого індивіда має бути щось об'єднуюче його часові етапи, завдяки чому він залишається незмінним. Для людини просторово-часова безперервність тіла забезпечує відповідний зв'язок; від дитинства до старості тіло прокладає безперервний шлях у світі, і саме цей шлях, а не будь-які індивідуальні особливості, з'єднує дитину та восьмидесятилітню людину. Це схоже на відому філософську проблему про корабель Тесея. Для таксона, ЯКЩО характеристики нерелевантні, ТОДІ це можуть бути лише родові зв'язки, які пов'язують девонський вид Rhyniognatha hirsti з сучасним метеликом-монархом як представниками таксону Insecta, розділених 400 млн років. Опозиційна позиція щодо цього ставить під сумнів передумову цього силогізму та стверджує, з епістемологічної точки зору, що члени таксонів лише емпірично розпізнаються на основі їхніх характеристик, а гіпотези спільного походження є результатами систематики, а не апріорними передумовами. Якщо немає ознак для визначення приналежності скам'янілості до таксономічної групи, то це лише некласифікований шматок породи.
Якщо походження є достатнім для безперервності таксону, то всі нащадки члена таксону також будуть включені в таксон, тому всі добросовісні таксони є монофілетичними; назви парафілетичних груп не заслуговують офіційного визнання. Оскільки «Пелікозаври» відносяться до парафілетичної групи, яка включає деяких чотириногих молюсків пермського періоду, але не їхніх існуючих нащадків, це не може бути дійсною назвою таксону. Водночас, не заперечуючи думку щодо назв лише монофілетичних груп, систематики-емпірики протистоять цьому есенціалізму предків, вказуючи на те, що пелікозаврів визнають парафілетичними через комбінацію синапоморфій і симплезіоморфій, тобто деякі з них ближче до ссавців, ніж до інших пелікозаврів. Матеріальне існування сукупності скам'янілостей і її статус як клади — це не одне й те саме. Монофілетичні групи заслуговують на увагу та назви, оскільки вони мають спільні цікаві властивості — синапоморфії — які є доказом, який дозволяє зробити висновок про спільне походження.
Історія
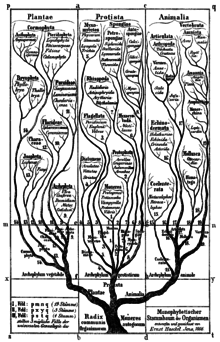
Філогенетична номенклатура є семантичним розширенням загального визнання розгалуження в ході еволюції, представленого на діаграмах Жана-Батіста Ламарка та інших вчених, таких як Чарльз Дарвін та Ернст Геккель. У 1866 році Е. Геккель вперше побудував єдине дерево всього живого на основі прийнятої на той час класифікації життя, яка базувалася на рангах та не містила таксонів, які Е. Геккель вважав поліфілетичними. У цьому дереві Геккель ввів ранг типу, який несе в собі конотацію монофілії в своїй назві (буквально означає «стебло»).
Відтоді точаться дискусії, як і якою мірою розуміння філогенії життя може використовуватися як основа для його класифікації, з поглядами, що варіюються від «числової таксономії» (фенетика) до " еволюційної систематики « (градистика) до „філогенетичної систематики“. Починаючи з 1960-х років, час від часу виникали безрангові класифікації, але загалом принципи та спільна мова традиційної номенклатури використовувалися всіма трьома вищенаведеними школами.
Більшість основних положень філогенетичної номенклатури (відсутність обов'язкових рангів і дещо близьке до філогенетичних визначень) можна простежити до 1916 року. Едвін Гудріч тоді інтерпретував назву Завропсиди, встановлену Т. Г. Хакслі 40 роками раніше, що включала птахів (Aves) та частину Reptilia, і придумав нову назву Theropsida, щоб включити ссавців, а також іншу частину Reptilia. Оскільки ці таксони існують вище рівнів, традиційно керованих правилами зоологічної номенклатури, Е. Гудріч не наполягав на рангах, але він чітко обмежив діагностичні ознаки, необхідні для розпізнавання та класифікації скам'янілостей, що належать до різних груп. Наприклад, стосовно п'ятої кістки плесни задньої кінцівки він написав, що „факти підтверджують нашу думку, оскільки ці ранні рептилії мають нормальні кістки плесни, як і їхні предки-амфібії. Отже, зрозуміло, що тут ми маємо цінну підтверджуючу рису, яка допоможе нам вирішити, чи належить даний вид до лінії еволюції Теропсидів чи Завропсидів“. Гудріч завершив свою статтю словами: „Присутність цих ознак показує, що всі живі рептилії належать до групи завропсидових, тоді як структура стопи дозволяє нам визначити спорідненість багатьох неповністю відомих викопних родів і зробити висновок, що лише певні вимерлі порядки можуть належать до гілки Теропсиди“. Гудріч висловив думку, що від назви Reptilia слід відмовитися після того, як філогенія рептилій стане краще вивченою.
Принцип, що лише клади повинні мати офіційні назви, став популярним у деяких колах у другій половині 20 століття. Цей принцип набув поширення разом із методами виявлення клад (кладистика) і був складовою частиною філогенетичної систематики (див. вище). Водночас стало очевидним, що обов'язкові ранги, які є частиною традиційних систем номенклатури, створюють проблеми. Деякі автори пропонували відмовитися від них взагалі, починаючи з відмови Віллі Генніга від його попередньої пропозиції визначити ранги як класи геологічного віку.
Перше опубліковане використання філогенетичної номенклатури датується 1986 роком. Незабаром з'явилися теоретичні статті про принципи філогенетичної номенклатури, а згодом — про її застосування (переважно до хребетних) (див. розділ „Література“).
З метою уникнення розколу в товаристві систематиків, Готьє запропонував двом членам ICZN застосовувати формальні таксономічні назви, керовані зоологічним кодексом, лише до клад (принаймні для надвидових таксонів) і відмовитися від рангів Ліннея, але вони члени негайно відкинули ці ідеї». Передумова назв у традиційній номенклатурі базується на типових зразках, а опис груп вважається таксономічним вибором, зробленим систематиками, які працюють над окремими групами, а не номенклатурним рішенням, прийнятим на основі апріорних правил у кодексах біологічної номенклатури. Бажання включити таксономічну об'ємність в номенклатурні визначення спонукало Kevin de Queiroz та ботаніка Philip Cantino до розробки власного кодексу номенклатури, PhyloCode, щоб врегулювати філогенетичну номенклатуру.
Альтернативна думка
Передова стаття Віллі Генніга спровокувала жваву дискусію щодо відносних переваг філогенетичної номенклатури проти таксономії Ліннея або пов'язаного підходу еволюційної таксономії, яка триває до сьогодні Деякі дебати, в яких брали участь кладисти, тривали з 19 століття. В. Генніг наполягав на користі різних схем класифікації , проте надавав перевагу своїй власній, стверджуючи, що категорії його системи мають «індивідуальність і реальність» на відміну від «позачасових абстракцій» класифікацій, заснованих на морфології.
Кажуть, що формальні класифікації, засновані на кладистичних міркуваннях, підкреслюють походження за рахунок описових характеристик. Тим не менш, більшість систематиків сьогодні по можливості уникають парафілетичних груп у систематиці Ліннея, а поліфілетичні таксони давно застарілі. Багато кладистів вважають, що традиційні кодекси зоологічної та ботанічної номенклатури повністю сумісні з кладистичними підходами, тому немає потреби заново винаходити систему назв, яка добре функціонувала протягом 250 років.
Міжнародний кодекс філогенетичної номенклатури
ICPN, або PhyloCode, — це сукупність правил і рекомендацій щодо філогенетичної номенклатури.
- ICPN регулює лише назви клад . Назви видів базуються на правилах традиційних кодів номенклатури .
- Принцип Пріоритету (або «пріоритет») застосовується для імен і визначень у ICPN. Точкою відліку для пріоритету було 30 квітня 2020 року.
- Визначення існуючих назв і нових імен разом із їхніми визначеннями потрібно публікувати в рецензованих роботах (відразу або після дати початку вживання) і реєструвати в онлайн-базі даних, після чого їх вважають дійсними.
Кількість прихильників широкого впровадження PhyloCode все ще невелика, і невідомо, наскільки широко його дотримуватимуться в майбутньому.
Список літератури
Джерела
- Hennig, Willi (1966). Phylogenetic systematics. Urbana, IL: Univ. of Illinois Press. с. 9. ISBN 978-0-252-06814-0. (reprinted 1979 and 1999)
Подальше читання
Нижче наведено кілька публікацій, не цитованих у списку літератури. Вичерпний перелік публікацій про філогенетичну номенклатуру можна знайти на веб -сайті Міжнародного товариства філогенетичної номенклатури .
- Bryant, Harold N. (1994). Comments on the phylogenetic definition of taxon names and conventions regarding the naming of crown clades. Syst. Biol. 43: 124–129. doi:10.1093/sysbio/43.1.124.
- Cantino, Philip D.; Olmstead, Richard G. (2008). Application of phylogenetically defined names does not require that every specifier be present on a tree. Syst. Biol. 57 (1): 157–160. PMID 18300028. doi:10.1080/10635150701883873.
- de Queiroz, Kevin (1992). Phylogenetic definitions and taxonomic philosophy. Biol. Philos. 7:295–313.
- Gauthier, Jacques A., Arnold G. Kluge, and Timothy Rowe (1988). The early evolution of the Amniota. Pages 103—155 in Michael J. Benton (ed.): The Phylogeny and Classification of the Tetrapods, Volume 1: Amphibians, Reptiles, Birds. Syst. Ass. Spec. Vol. 35A. Clarendon Press, Oxford.
- Gauthier, Jacques, David Cannatella, Kevin de Queiroz, Arnold G. Kluge, and Timothy Rowe (1989). Tetrapod phylogeny. Pages 337—353 in B. Fernholm, K. Bremer, and H. Jörnvall (eds.): The Hierarchy of Life. Elsevier Science B. V. (Biomedical Division), New York.
- Ghiselin, M. T. (1984). Definition," "character," and other equivocal terms. Syst. Zool. 33 (1): 104–110. JSTOR 2413135. doi:10.2307/2413135.
- Keesey, T. Michael (2007). A mathematical approach to defining clade names, with potential applications to computer storage and processing. Zool. Scr. 36 (6): 607–621. doi:10.1111/j.1463-6409.2007.00302.x.
- Laurin, Michel (2005). The advantages of phylogenetic nomenclature over Linnean nomenclature. Pages 67–97 in A. Minelli, G. Ortalli, and G. Sanga (eds): Animal Names. Instituto Veneto di Scienze, Lettere ed Arti; Venice.
- Lee, Michael S. Y. (2005). Choosing reference taxa in phylogenetic nomenclature. Zool. Scr. 34 (3): 329–331. doi:10.1111/j.1463-6409.2005.00196.x.
- Rowe, Timothy (1987). Definition and diagnosis in the phylogenetic system. Syst. Zool. 36 (2): 208–211. JSTOR 2413270. doi:10.2307/2413270.
- Rowe, Timothy; Gauthier, Jacques (1992). Ancestry, paleontology and definition of the name Mammalia. Syst. Biol. 41 (3): 372–378. doi:10.1093/sysbio/41.3.372.
- Sereno, Paul C. (1998). A rationale for phylogenetic definitions, with application to the higher-level taxonomy of Dinosauria. Neues Jahrbuch für Geologie und Paläontologie - Abhandlungen 210: 41–83. doi:10.1127/njgpa/210/1998/41.
- Sereno, Paul C. (1999). Definitions in phylogenetic taxonomy: critique and rationale. Syst. Biol. 48 (2): 329–351. PMID 12066711. doi:10.1080/106351599260328.
- Sereno, Paul C. (2005). The Logical Basis of Phylogenetic Taxonomy. Syst. Biol. 54 (4): 595–619. PMID 16109704. doi:10.1080/106351591007453.
- Taylor, Michael P. (2007). Phylogenetic definitions in the pre-PhyloCode era; implications for naming clades under the PhyloCode. PaleoBios 27: 1–6.
- Wilkinson, Mark (2006). Identifying stable reference taxa for phylogenetic nomenclature. Zool. Scr. 35: 109–112. doi:10.1111/j.1463-6409.2005.00213.x.
- Wyss, A. R.; Meng, J. (1996). Application of phylogenetic taxonomy to poorly resolved crown clades: a stem-modified node-based definition of Rodentia. Syst. Biol. 45 (4): 559–568. doi:10.1093/sysbio/45.4.559.
|
| |||||||||||||||||||||||