
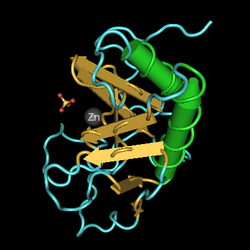
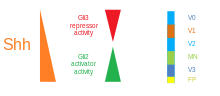
Sonic hedgehog (SHH) — це один з трьох білків сигнальної трансдукції ссавців, а також ген, що його кодує. Інші два — desert hedgehog (DHH) та Indian hedgehog (IHH). Всіх їх називають hh-гомологи. SHH є найкраще вивченим лігандом сигнальної трансдукції. Він відіграє ключову роль у регулюванні органогенезу хребетних, наприклад, в утворенні пальців на кінцівках та формуванні мозку. Sonic hedgehog — найкращий приклад морфогену, який визначається моделлю трикольорового прапору Льюїса Вольперта — молекули, яка дифундує, утворюючи градієнт концентрації та має різний вплив на клітини розвиваючогося ембріона в залежності від його концентрації. SHH залишається важливим білком у організмі дорослої людини. Він контролює клітинний поділ дорослих стовбурових клітин і бере участь у розвитку деяких видів раку.
Sonic hedgehog — білок, який кодується однойменним геном, розташованим у людей на короткому плечі 7-ї хромосоми. Як білок, так і ген, що цього кодує, можна також позначати абревіатурою Shh
Довжина поліпептидного ланцюга білка становить 462 амінокислот, а молекулярна маса — 49 607.
| 10 | 20 | 30 | 40 | 50 | ||||
|---|---|---|---|---|---|---|---|---|
| MLLLARCLLL | VLVSSLLVCS | GLACGPGRGF | GKRRHPKKLT | PLAYKQFIPN | ||||
| VAEKTLGASG | RYEGKISRNS | ERFKELTPNY | NPDIIFKDEE | NTGADRLMTQ | ||||
| RCKDKLNALA | ISVMNQWPGV | KLRVTEGWDE | DGHHSEESLH | YEGRAVDITT | ||||
| SDRDRSKYGM | LARLAVEAGF | DWVYYESKAH | IHCSVKAENS | VAAKSGGCFP | ||||
| GSATVHLEQG | GTKLVKDLSP | GDRVLAADDQ | GRLLYSDFLT | FLDRDDGAKK | ||||
| VFYVIETREP | RERLLLTAAH | LLFVAPHNDS | ATGEPEASSG | SGPPSGGALG | ||||
| PRALFASRVR | PGQRVYVVAE | RDGDRRLLPA | AVHSVTLSEE | AAGAYAPLTA | ||||
| QGTILINRVL | ASCYAVIEEH | SWAHRAFAPF | RLAHALLAAL | APARTDRGGD | ||||
| SGGGDRGGGG | GRVALTAPGA | ADAPGAGATA | GIHWYSQLLY | QIGTWLLDSE | ||||
| ALHPLGMAVK | SS |
Кодований геном білок за функціями належить до гідролаз, протеаз, білків розвитку. Білок має сайт для зв'язування з іонами металів, іоном цинку, іоном кальцію. Локалізований у клітинній мембрані. Також секретований назовні.
Відкриття та назва
Ген sonic hedgehog (hh) вперше був ідентифікований у плодової мушки Drosophila melanogaster вченими англ. Christiane Nüsslein-Volhard та англ. Eric Wieschaus, та опублікований в 1980 році. , за що вони отримали Нобелівську премію в 1995 році разом з генетиком Едвардом Б. Льюїсом, який визначив гени, що контролюють патерни сегментації ембріонів дрозофіли. Втрата функціонування hh у мутантного фенотипу призводить до того, що ембріони покриваються зубцями, невеликими точковими виступами, що нагадують голки їжака.
Дослідження, спрямовані на пошук еквівалента sonic hadgehog у хребетних, проведені Філіпом Інгегом, Ендрю П. МакМахоном та Кліффордом Табіном, показали три подібні гени. Два з них, desert hedgehog та Indian hedgehog — були названі на честь видів їжаків, а sonic hadgehog був названий на честь популярного в той час персонажа відеогри SEGA, їжака Соніка. Назва була обрана Робертом Ріддлом, постдоктором лабораторії Tabin, після того, як він побачив комікс з даним персонажем, зроблений його дочкою з Англії.
Функції
З hh-гомологів було виявлено, що SHH має найважливішу роль у розвитку, виступаючи в ролі морфогену, що залучає до складання множинних систем, включаючи кінцівку та серединну структуру головного мозку, спинного мозку,Таламусу в zona limitans intrathalamica, легенів, і зубів.
Мутації в гені SHH людини, викликають голопрозенцефалію третього типу внаслідок втрати вентральної середньої лінії. SHH виділяється в зоні поляризаційної активності, яка розташована на задній стороні долоні кінцівок у ембріоні. Шлях транскрипції SHH також був пов'язаний з утворенням специфічних видів ракових пухлин, у тому числі ембріональної пухлини мозочка та медулобластоми , а також прогресуванням пухлин раку передміхурової залози . Для того, щоб SHH був виражений у кінцівках ембріонів, що розвиваються, морфоген, що називається фактором росту фібробластів, повинен виділятися з верхівкового ектодермального гребеню.

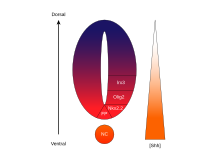
SHH також показав, що він діє як аксональний навідник. Показано, що SHH направляє комісулярні аксони на вентральну середню лінію спинного мозку, що розвивається. Зокрема, SHH притягує аксони нейронів гангліїв сітківки (RGC) у низьких концентраціях і відштовхує їх у більш високих концентраціях. Показано, що відсутність SHH забезпечує зародження недорозвинених задніх кінцівок у китоподібних (кити та дельфіни).
Формування центральної нервової системи
Молекула сигнального білку Sonic hedgehog (SHH) відіграє різні ролі при формуванні центральної нервової системи (ЦНС) під час розвитку хребетних. Одним з найбільш характерних функцій SSH є його роль у індукції вентральної пластинки та різноманітних типів вентральних клітин у нервовій трубці . Хорда, структура, сформована з осьової мезодерми, виробляє SHH, який міжклітинно проходить до вентральної ділянки нервової трубки, і сигналізує цим клітинам утворювати вентральну пластинку. Інша точка зору щодо індукції вентральної пластинки припускає, що деякі клітини-попередники, розташовані в хорді, мігрують в нервову пластину до її утворення, згодом породжуючи вентральну пластинку.
Сама нервова трубка є попередником хребетної ЦНС, а вентральна пластинка є спеціалізованою структурою і розташована у середній точці нейронної трубки. Докази, що підтверджують діяльність хорди як центру сигналізації походять від досліджень, в яких другий нотохорд імплантується поблизу нервової трубки in vivo, що призводить до утворення вентральної пластинки в межах нервової трубки .
SHH - це протеїн, який опосередковує сигнальну діяльність нотохорду та поверальної пластинки. Дослідження, що вивчають експресію SHH in vitro та in vivo , проказують, що вона призводить до індукції вентральної пластинки та диференціювання моторних нейронів та вентральних інтернейронів. З іншого боку, мишей-мутантів по SHH відсутні вентральні характеристики спинного мозку . In vitro блокування SHH сигналізації з використанням антитіл проти нього проявляє подібні фенотипи SHH exerts its effects in a concentration-dependent manner,. Вплив SHH має залежність від концентрації, тому висока концентрація SHH призводить до локального гальмування клітинної проліферації . Це пригнічення призводить до того, що вентральна пластинка стає тонкою в порівнянні з латеральними областями нервової трубки. Зниження концентрації SHH призводить до проліферації клітин та індукції різних типів вентральних нейронних клітин. Після того, як сформовано вентральну пластинку, клітини, що знаходяться у цьому регіоні, згодом експресують SHH , що генерує градієнт концентрації в нервовій трубці .
Морфогенетична активність
Концентрація та залежна від часу детермінуюча активність SHH у вентральній нервовій трубці, робить його головним прикладом морфогену. У хребетних сигналізація SHH у вентральній частині нейронної трубки найчастіше відповідальна за індукцію вентральної пластинки клітин та моторних нейронів. SHH виходить з хорди та вентральної пластинки нервової трубки, що розвивається, щоб створити градієнт концентрації, який охоплює дорзо-вентральну вісь . Більш високі концентрації ліганду SHH зустрічаються найбільш вентральних частинах нервової трубки і хорди, в той час як менші концентрації зустрічаються в більш дорсальних ділянках нервової трубки . Градієнт концентрації SHH був візуалізований у нервовій трубці мишей, модифікованих для експресії комплексу білків SHH — GFP, щоб візуалізувати цей градуйований розподіл SHH під час формування вентральної нервової трубки.
Формування зубів
SHH - сигнальна молекула, кодована однойменним геном. SHH відіграє дуже важливу роль в органогенезі і, головне, розвитку черепно-лицьового відділу. SHH є сигнальною молекулою, що в першу чергу дифундує вздовж градієнта концентрації, що чинить різний вплив на віддалені одна відносно одної клітини. У ранньому розвитку зубів SHH вивільняється з первинного емалевого вузла, сигнального центру, що забезпечує позиційну інформацію у розвитку зубів та регуляції росту зуба . Зокрема, SHH необхідний для росту епітеліальних шийних петель, де зовнішні та внутрішні епітелії приєднуються і утворюють резервуар для стовбурових клітин пульпи зуба. Після того як первинні емалеві вузли апоптозуються, утворюються вторинні емалеві вузли. Первинні емалеві вузли виділяють SHH у поєднанні з іншими сигнальними молекулами, щоб згущати оральний ектодерм і починати формувати складні форми коронки зуба під час диференціації та мінералізації . В моделі нокаутного гена відсутність SHH свідчить про голопросенцефалію. Крім того, SHH активує молекули Gli2 і Gli3. Мутантні ембріони по Gli2 та Gli3 мають ненормальний розвиток різців, а також дрібних молярів .
Процесинг
SHH проходить ряд етапів обробки, перш ніж він виділяється з клітини. Щойно синтезований SHH важить 45 кДа. Він містить коротку сигнальну послідовність на її N-кінці, яка визнає розпізнавання білку під час транслокації в ендоплазматичний ретикулум. Після завершення транслокації сигнальна послідовність видаляється сигнальною пептидазою в ендоплазматичному ретикулумі. Там SHH проходить автопроцесинг і генерує N-кінцевий сигнальний домен 20 кДа (SHH-N) і 25-кДа C-кінцевий сигнальний домен невідомого призначення. Розщеплення каталізується протеазою в межах С-кінцевого домену. Під час реакції розщеплення молекула холестеролу приєднується до С-кінця SHH-N. Таким чином, C-кінцевий домен діє як інтеїн та холестеролова трансфераза.

Інший гідрофобний фрагмент, пальмітат, приєднується до альфа-аміну N-кінцевого цистеїну SHH-N. Ця модифікація необхідна для ефективної сигналізації, і приводить до 30-кратного збільшення потенції над непальмітильованою формою і виконується Protein-cysteine N-palmitoyltransferase, членом сімейства мембран-ассоційованих О-ацетилтрансфераз.
Роботнікінін
Потенційний інгібітор SHH сигналіхації отримав назву роботнікінін, на честь героя-антагоніста Sonic The Hedgehog, Доктора Айво Роботніка.
Суперечка щодо назви
Ген sonic hadgehog пов'язаний з патологією голопрозенцефалією, яка призводить до серйозних дефектів головного мозку, черепа та обличчя. Цей факт закликає клініцистів та вчених критикувати назву на підставі того, що в такому контексті вона звучить надто легковажно. Ще менш комічна ситуація виникає коли пацієнтам або батькам пацієнтів з серйозним розладом сповіщають, що вони або їхні діти мають мутацію у гені "Sonic Hadgehog", названому ім'ям персонажу дитячої відеогри.
Галерея



Література
- Norbnop P., Srichomthong C., Suphapeetiporn K., Shotelersuk V. (2014). ZRS 406A>G mutation in patients with tibial hypoplasia, polydactyly and triphalangeal first fingers.. J. Hum. Genet. 59: 467 — 470. PubMed DOI:10.1038/jhg.2014.50
- El-Jaick K.B., Brunoni D., Castilla E.E., Moreira M.A., Orioli I.M. (2005). SHH Ile111Asp in alobar holoprosencephaly in a proposita, whose mother had only a solitary median maxillary incisor.. Am. J. Med. Genet. A 136: 345 — 345. PubMed DOI:10.1002/ajmg.a.30624
- Ribeiro L.A., Richieri-Costa A. (2005). Single median maxillary central incisor, hypophyseal tumor, and SHH mutation.. Am. J. Med. Genet. A 136: 346 — 347. PubMed DOI:10.1002/ajmg.a.30625
- Maity T., Fuse N., Beachy P.A. (2005). Molecular mechanisms of Sonic hedgehog mutant effects in holoprosencephaly.. Proc. Natl. Acad. Sci. U.S.A. 102: 17026 — 17031. PubMed DOI:10.1073/pnas.0507848102
- Richieri-Costa A., Ribeiro L.A. (2006). Holoprosencephaly-like phenotype: clinical and genetic perspectives.. Am. J. Med. Genet. A 140: 2587 — 2593. PubMed DOI:10.1002/ajmg.a.31378
- Tate G., Kishimoto K., Mitsuya T. (2000). Expression of Sonic hedgehog and its receptor Patched/Smoothened in human cancer cell lines and embryonic organs.. J. Biochem. Mol. Biol. Biophys. 4: 27 — 34.