
|
Археї (Archaea) Час існування: 3.5–0 Ga Палеоархей чи можливо Еоархей – т. ч. | ||||||||
|---|---|---|---|---|---|---|---|---|
 Halobacteria sp. штам NRC-1, довжина кожної клітини близько 5 μm
| ||||||||
| Біологічна класифікація | ||||||||
| ||||||||
|
Царства
| ||||||||
| ||||||||
|
Посилання
| ||||||||
|
Архе́ї (лат. Archaea, від грец. αρχαία — «старі», також архебактерії — Archaebacteria) — одна з груп живих організмів, до якої належать мікроскопічні одноклітинні прокаріоти, що дуже відрізняються низкою фізіолого-біохімічних ознак від справжніх бактерій (еубактерій). Хоча досі є невизначеність в точному філогенезі цих груп, археї, еукаріоти і бактерії є фундаментальними групами в так званій системі трьох доменів. Подібно до бактерій, археї — це одноклітинні організми, що не мають ядра і тому класифікуються як прокаріоти (Prokaryota) — відомі також як Monera в таксономії п'яти царств. Археїв спочатку було виявлено в екстремальних середовищах, але потім їх було знайдено в усіх типах екосистем. Архебактерії суттєво відрізняються від інших мікроорганізмів за складом і послідовністю нуклеотидів у рибосомних і транспортних РНК.
Історія
Археїв було ідентифіковано 1977 року Карлом Воузом (Carl Woese) і Джорджем Фоксом (George Fox), засновуючись на їхній відмінності від інших прокаріотів (філогенетичні дерева, що базуються на 16S рРНК). Коли 1977 року біолог Карл Воуз вивчав генетику мікробів, що виробляли метан, він зрозумів, що вони відрізняються від усіх відомих бактерій. Стінка клітини була унікальною — виробляла унікальні ензими і її генетичний ряд не був схожий ні на що досліджуване раніше. Дуже швидко — протягом години — стало зрозуміло, що є ще щось, третя група. Це був момент відкриття. Карл Воуз відкрив третю форму життя — групу одноклітинних, яку він назвав археями. Він відкрив форму життя, яка здатна існувати будь-де, включаючи найнеймовірніші місця. Цю групу було спочатку названо археобактеріями (Archaebacteria) на відміну від еубактерій (Eubacteria), і вони розглядалися як царства або підцарства. Воуз переконував, що археї є представниками зовсім іншої гілки живих істот. Він пізніше перейменував групи в архей та бактерій, щоб підкреслити це, і аргументував тим, що разом з еукаріотами вони охоплюють три групи живих організмів.
Біологічний термін архея (археї) не слід плутати з Архейською геологічною ерою, також відомою як Археозойська ера. Останній термін посилається на стародавній період земної історії, коли археї та бактерії були єдиними клітинними організмами, що жили на планеті. Імовірні скам'янілості цих мікробів були датовані майже 3,8 мільярдів років тому.
Археї, бактерії та еукаріоти

Археї подібні до інших прокаріотів у більшості аспектів структури клітини та метаболізму. Проте, їхня генетична транскрипція і трансляція — два центральні процеси в молекулярній біології — не виявляють типових для бактерій особливостей, але надзвичайно подібні до цих процесів в еукаріотах. Наприклад, система трансляції архей використовує еукаріотичні фактори ініціації та елонгації, а їх система транскрипції застосовує TATA-зв'язувані білки і TFIIB, як в еукаріотах. Гени тРНК та рРНК археї містять унікальні археальні інтрони, які не подібні ні до еукаріотних, ні до бактеріальних інтронів.
Деякі інші характеристики також виділяють архей. Подібно до бактерій і еукаріотів, археї мають засновані на гліцерині фосфоліпіди. Проте, три особливості ліпідів архей незвичайні:
- Ліпіди архей унікальні, тому що стереохімія гліцерину — обернена до тієї, що знайдена в бактеріях і еукаріотах. Це переконливе свідчення іншого біосинтетичного шляху.
- Більшість бактерій і еукаріотів мають мембрани, складені переважно з естер-ліпідів гліцерину, тоді як археї мають мембрани, складені з етер-ліпідів гліцерину. Навіть, коли бактерії мають етер-зв'язані ліпіди, стереохімія гліцерину — бактерійна форма. Ці відмінності, можливо, є адаптацією частини архей до гіпертермофілії. Проте, варто відзначити, що навіть мезофілічні археї мають етер-зв'язані ліпіди.
- Ліпіди архей засновані на ізопреноїдному боковому ланцюжку. Це п'ятивуглецева хімічна група, також звичайна для каучуку і є компонентом деяких вітамінів, поширена в бактеріях і еукаріотах. Проте, тільки археї застосовують ці речовини в своїх клітинних ліпідах, часто як бокові ланцюжки C-20 (чотири мономера) або C-40 (вісім мономерів). У деяких архей ізопреноїдний боковий ланцюжок C-40 — фактично достатньо довгий, щоб пройти крізь мембрану, формуючи моношар замість клітинної мембрани з групами фосфату гліцерину на обох кінцях.
Ця адаптація найпоширеніша в надзвичайно теплолюбивих археях. Хоча вони і не унікальні, клітинні стінки архей також незвичні. Наприклад, клітинні стінки більшості архей утворені зовнішніми шарами білків або S-шаром. S-шари розповсюджені в бактеріях, де вони слугують єдиним компонентом клітинної стінки в деяких організмах (наприклад, у Planctomyces) або зовнішній шар в багатьох організмах з пептидогліканами. За винятком однієї групи метаногенів, археї не мають пептидогліканової стінки. Однак навіть у цьому випадку, пептидоглікани дуже відрізняються від різновиду, знайденого в бактеріях. Археї також мають джгутики, які значно відрізняються за складом від дуже подібних джгутиків бактерій. Бактерійні джгутики — це змінена система секреції III типу, тоді як джгутики архей нагадують ворсинки IV типу (type IV pili), які використовують sec-залежну систему секреції, дещо подібну, але все ж відмінну від системи секреції II типу.
Середовище

Багато архей — екстремофіли. Деякі живуть при дуже високих температурах, часто вище 100 °C, як ті, котрих знайшли в гейзерах і чорних курцях. Інших знайдено в дуже холодних середовищах або в надзвичайно солоній, кислій, або лужній воді. Проте, деякі археї — мезофіли, живуть у середовищах, подібних до болота, стічних вод і ґрунту. Багато метаногенних архей знайдено в травних трактах тварин, наприклад, жуйних тварин, термітів і людей. Археї не патогенні, і невідомо, щоб які-небудь з них викликали хворобу.
Архей звичайно поділяють на три групи, відповідно до середовища. Це — галофіти, метаногени і термофіли. Галофіли живуть у надзвичайно солоних навколишніх середовищах (наприклад, багато з них живуть у Мертвому морі та на півдні затоки Сан-Франциско, надаючи їй яскравих кольорів: від червоного до зеленого). Метаногени живуть в анаеробних навколишніх середовищах і виробляють метан. Їх можна знайти в осадах або в кишечниках тварин. Термофіли живуть у місцях з високими температурами, як, наприклад, гарячі джерела. Ці групи не обов'язково узгоджуються з молекулярним філогенезом, не повні та не взаємовиключні. Тим не менш, вони — корисний початковий пункт для докладнішого вивчення.
Нещодавно декілька досліджень показали[джерело?], що археї існують не тільки в мезофільних та термофільних навколишніх середовищах, але також іноді присутні у значній кількості при низьких температурах. Стає все більш прийнятим, що метаногени зазвичай присутні в низькотемпературних середовищах, як, наприклад, холодні опади. Деякі дослідження навіть свідчили, що за цих температур шлях метаногенезу може змінюватися завдяки термодинамічним обмеженням, зумовленим низькими температурами. Можливо, навіть, важливішим є те, що велику кількість архей знайдено у більшості океанів, у холодних середовищах (Giovannoni і Stingl, 2005). Ці археї, які належать до груп, раніше не досліджених, можуть траплятися в надзвичайно високих кількостях (до 40% мікробної біомаси), хоча вони не були ізольовані в чистій культурі. Зараз ми не маємо майже ніякої інформації щодо фізіології цих організмів, що означає, що їх вплив на глобальні біогеохімічні цикли невідомий. Одне недавнє дослідження (Könneke й інші, 2006) показало, проте, що одна група морських Crenarchaeota здатна до фіксації азоту, а це — риса, досі невідома серед архей.
Генетика
Як правило, археї мають одиночну кільцеву хромосому, розмір якої може сягати 5 751 492 пар нуклеотидів у Methanosarcina acetivorans, що має найбільший відомий геном серед архей. Одну десяту розміру цього геному складає геном з 490 885 парами нуклеотидів у Nanoarchaeum equitans, що має найменший геном серед архей; він містить лише 537 генів, які кодують білки. Також у архей виявлені дрібніші незалежні молекули ДНК, так звані плазміди. Ймовірно, плазміди можуть передаватися між клітинами при фізичному контакті, під час процесу схожого на кон'югацію бактерій.
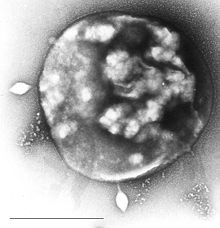
Археї можуть бути вражені вірусами, що містять дволанцюжкову ДНК. Ці види не споріднені з іншими групами вірусів і мають різноманітні незвичайні форми, так як пляшки, гачки, краплі тощо. Ці віруси були ретельно вивчені на термофілах, в основному рядів Sulfolobales і Thermoproteales. У 2009 році був відкритий вірус, що містить одноланцюжкову ДНК і вражає галофільні археї. Захисні реакції архей проти вірусів можуть включати механізми близький до РНК-інтерференції еукаріот.
Археї генетично відмінні від еукаріот і бактерій, причому до 15 % білків, що кодуються одним геном бактерії, є унікальними для цього домену, хоча функції багатьох цих білків невідомі. Більша частина унікальних білків, функція яких відома, належить евріархеотам і задіяна в метаногенезі. Білки, спільні для архей, бактерій та еукаріот, беруть участь в основних клітинних функціях і стосуються в основному транскрипції, трансляції і метаболізму нуклеотидів. До інших відмінностей архей можна віднести організацію генів, що виконують пов'язані функції (приміром, гени, що відповідають за різні етапи одного й того ж метаболічного процесу), в оперони і великі відмінності у будові тРНК і їх аміноацил-тРНК синтетаз.
Транскрипція і трансляція архей більше нагадують ці процеси в клітинах еукаріот, ніж бактерій, причому РНК-полімераза і рибосоми архей дуже близькі до аналогічних структур у еукаріот. Хоча у архей є лише один тип РНК-полімерази, за будовою і функцією вона близька до РНК-полімерази II еукаріот, при цьому подібні групи білків (головні фактори транскрипції) забезпечують зв'язування РНК-полімерази з промотором гена. у той же час інші фактори транскрипції архей більш близькі до таких у бактерій. Процесинг РНК у архей простіше, ніж у еукаріот, оскільки більшість генів не містить інтронів, хоча в генах їх тРНК і рРНК їх достатньо багато, також вони присутні у невеликій кількості генів, що кодують білки.
Розмноження
Археї розмножуються безстатевим шляхом: бінарним чи множинним поділом, фрагментацією чи брунькуванням. Мейозу не відбувається, тому навіть якщо представники конкретного виду архей існують у більш ніж одній формі, всі вони мають однаковий генетичний матеріал. Клітинний поділ визначається клітинним циклом: після того, як хромосома реплікована і дві дочірні хромосоми розійшлись, клітина ділиться. Деталі були вивчені лише для роду Sulfolobus, але особливості його циклу дуже схожі з особливостями у еукаріот і у бактерій. Реплікація хромосом починається з багатьох точок початку реплікації за допомогою ДНК-полімерази, схожої на аналогічні ферменти еукаріот. Однак білки, що керують клітинним поділом, такі як FtsZ, які формують стискаюче кільце навколо клітини, і компоненти септи, що проходить через центр клітини, схожі з їх бактеріальними еквівалентами.
Археї не утворюють спори. Деякі види Haloarchaea можуть зазнавати зміни фенотипу й існувати як клітини декількох різних типів, включно з товстостінними клітинами, стійкими до осмотичного шоку, що дозволяють археям виживати у воді з низькою концентрацією солі. Однак ці структури не слугують для розмноження, а скоріш допомагають археям освоювати нові середовища проживання.
Еволюція і класифікація
Назва говорить про те, що ці організми в наш час[коли?] багатьма дослідниками вважаються найдавнішими живими організмами на Землі.
Археї поділено на дві основні групи, засновані на деревах 16S рРНК: Euryarchaeota і Crenarchaeota. Дві інші групи було також створено для певних екологічних зразків і особливого виду Nanoarchaeum equitans, знайдених у 2002 Карлом Стеттером (Karl Stetter), але їх подібність до інших груп все ще досліджено дуже погано.
Карл Воуз (Carl Woese) переконав, що бактерії, археї і еукаріоти являють собою унікальні спадкові лінії, які досить рано відхилилися від спадкового прогенота (Progenote) з погано розвиненим генетичним апаратом. Ця гіпотеза відображена в імені археї (Archaea, від Грецького archae або стародавній). Пізніше він формально розглядав ці групи як імперії, кожна з яких охоплює декілька царств. Цей поділ став найпопулярнішим, хоча ідея прогенотів не всіма підтримується. Деякі біологи, проте, переконують, що, археобактерії і еукаріоти походять від спеціалізованих бактерій.
Взаємовідносини між археями та еукаріотами залишаються важливою проблемою. Враховуючи подібність, зазначену вище, багато генетичних дерев розташовують ці дві групи разом. Деякі ставлять еукаріотів ближче до Eurarchaeota, ніж до Crenarchaeota, хоча мембранна хімія пропонує навпаки. Проте відкриття архея-подібних генів у певних бактеріях, наприклад, Thermotoga, робить їх взаємовідношення важким для визначення. Деякі дослідники вважають, що еукаріоти виникли через змішення археї і бактерії, де вони стали ядром і цитоплазмою, що поясняє різну генетичну схожість, але ця теорія не може пояснити походження клітинних структур.
Внутрішня класифікація
Архебактерії різноманітні за типом обміну речовин, фізіологічними і екологічними особливостями: серед них зустрічаються аероби і анаероби, хемогетеротрофи і хемоавтотрофи, нейтрофіли і ацидофіли. Деяким архебактеріям (галобактеріям) притаманний особливий тип фотосинтезу, коли світло поглинається не хлорофілом, а бактеріородопсином. Іншим археям властивий енергетичний процес, в результаті якого утворюється метан. Описано понад 40 видів археїв (25 родів), що належать до 5 різних груп: метаногени, сіркоокислюючі термоацидофіли, сірковідновлюючі термофіли, галобактерії, термоплазми.
Розшифровка одиночних генів призвела до розшифровки цілих геномів, зараз 24 архейні геноми завершено і 22 частково завершено (1).
Джерела
- Howland, John L. (2000). The Surprising Archaea: Discovering Another Domain of Life. Oxford: Oxford University Press. ISBN 0-19-511183-4.
- Giovannoni, S.J. and Stingl, U. (2005). Molecular diversity and ecology of microbial plankton. Nature 437: 343–348.
- Könneke, M., Bernhard, A.E., de la Torre, J.R., Walker, C.B., Waterbury, J.B. and Stahl, D.A. (2005). Isolation of an autotrophic ammonia-oxidizing marine archaeon. Nature 437: 543–546.
- Lake, J.A. (1988). Origin of the eukaryotic nucleus determined by rate-invariant analysis of rRNA sequences. Nature 331: 184–186.
- Woese, Carl R.; Fox, George E. (1977). Phylogenetic Structure of the Prokaryotic Domain: The Primary Kingdoms. Proceedings of the National Academy of Sciences of the United States of America 74 (11): 5088-5090.
- Woese, Carl R., Kandler, Otto, Wheelis, Mark L (1990). Towards a natural system of organisms: Proposal for the domains Archaea, Bacteria, and Eucarya. Proceedings of the National Academy of Sciences 87 (12): 4576-4579.
- 100 найбільших відкриттів. Біологія (2005), Discovery
Посилання
- Хто такі Археї? [Архівовано 31 жовтня 2013 у Wayback Machine.] (укр.)
- Археї
- ArchaeaWeb — з UNSW — Інформація про архей
- Введення до архей, їх екології, систематики та морфології [Архівовано 29 жовтня 2004 у Wayback Machine.]
- Перспектива використування екстремофілів для антимікробних ліків, Др. Сара Малоні [Архівовано 23 квітня 2006 у Wayback Machine.] Цитата: «…Ground breaking research on extremophiles continues to this day, with the recently-discovered 22nd genetically-encoded amino acid — pyrrolysine — from the archaeon, Methanosarcina barkeri, (Hao et al., 2002; Srinivasan et al., 2002)…»
- Новини BBC 21 червня, 1999: Найскладніший організм розкриває генетичні секрети [Архівовано 5 лютого 2007 у Wayback Machine.] Цитата: «…It [Pyrococcus abyssi] likes conditions that the vast majority of other organisms would find impossible to live in. It thrives best at temperatures of about 103 degrees [Celsius] and under pressures of about 200 atmospheres….»
- Сторінка Pyrococcus abyssi на Genoscope [Архівовано 24 жовтня 2006 у Wayback Machine.]