
Співвідношення статей — відношення числа самців до числа самок у роздільностатевих популяціях.
Співвідношення статей поряд з статевим диморфізмом є важливою характеристикою роздільностатевих популяцій. Зазвичай його вказують за кількістю самців, що припадають на 100 самок, часткою чоловічих особин або у процентах. Залежно від стадії онтогенезу розрізняють первинне, вторинне і третинне співвідношення статей. Первинне — це співвідношення статей у зиготах після запліднення; вторинне — співвідношення статей при народженні і, нарешті, третинне — співвідношення статей зрілих, здатних розмножуватися особин популяції.
Підвищенне первинне і вторинне співвідношення статей
В даний час загальновизнано, що у більшості видів тварин і рослин основним механізмом, визначаючим стать, є хромосомний. Оскільки в ході гаметогенезу гамети, містять Х- або Y-хромосоми, виробляються в рівному числі, вважалося, що цей механізм забезпечує приблизно рівні частки статей при зачатті. Однак, вторинне співвідношення статей залежить не тільки від пропорції гетерогамет, але і від багатьох інших факторів. Наприклад, від співвідношення швидкостей старіння і елімінації Х- і Y-несучих сперміїв в організмі самця, від їхньої здатності досягти яйцеклітини і запліднити її, від спорідненості яйцеклітин до Х- або Y-сперміїв, нарешті, від життєздатності чоловічих і жіночих ембріонів на різних стадіях ембріонального розвитку.
Давно було помічено, що у багатьох видів тварин вторинне співвідношення статей дещо відрізняється від пропорції 1 : 1 в бік надлишку самців приблизно 105–106 самців на 100 самок. Найбільше надійних даних зібрано щодо людей. Середнє значення вторинного співвідношення статей по всіх людських популяціях становить близько 106. Урахування неоднакової (диференціальної) смертності статей під час ембріональної стадії розвитку, ще більше зрушує екстрапольоване значення первинного співвідношення статей від пропорції 1 : 1. Всі доступні дані щодо статевого складу викиднів і мертвонароджених у людини показують, що хлопчиків помирає під час утробного життя в 2-4 рази більше, ніж дівчаток. Таким чином, первинне співвідношення статей у людини, по всій видимості, відхиляється від пропорції 1: 1 в бік надлишку чоловічих зигот, і воно, мабуть, більше, ніж вторинне. У людини співвідношення статей при народженні може штучно порушуватися за рахунок абортів і дітовбивства,,.
Зв'язок між вторинним і третинним співвідношенням статей
Між вторинним і третинним співвідношенням статей існує пряма залежність — чим більше народжуваність чоловічих особин, тим більше їх може дожити до зрілого віку.
Багато даних свідчать також про те, вторинне співвідношення статей залежить від третинного. У восьми видів рослин і тварин (дрімота, гуппі, кліщі (3 види), дрозофіла, миші, пацюки, лісовий північноамериканський бабак і людина) прямими експериментами було показано, що збільшення третинного співвідношення статей призводить до зменшення вторинного.
Комахи
У комах (бджіл та інших перетинчастокрилих, червеців, кліщів) з запліднених яєць виходять самки (або самки і самці), а з незапліднених розвиваються тільки самці. Тому, чим менше самців в вихідній популяції, тим в середньому менше яєць піддається заплідненню і тим більше самців виходить у потомстві.


Людина
Статевий дисбаланс
Гендерний дисбаланс — демографічний ефект, що виникає в зв'язку з війнами, наприклад, в Європі після Першої світової війни і після Другої світової війни (у Росії, див. Демографія в Радянському Союзі), або як результат внутрішньої політики (напр., одна сім'я — одна дитина).
Люди не є строго моногамними, і у тих народів, у яких поширене багатожонство і гареми, виникають різні відхилення третинного співвідношення статей від пропорції 1 : 1, як правило у бік його зменшення. У нігерійців антрополог Норткот У. Томас помітив зростання вторинного співвідношення статей із збільшенням числа дружин.
Вторинне співвідношення статей у гаремах збільшується приблизно до 62% (гареми: імператора Чжу Юаньчжана (1328–1398, Китай) 26 хлопчиків: 16 дівчаток; фараона Рамзеса II (1317–1251 д.н. е., Єгипет) 74 дружини, 111 : 68; та султана Мауле Ісмаїла (1646–1727, Марокко) 548 : 340). До даних отриманих для країн, де більше цінується народження хлопчиків, слід ставитися з обережністю, беручи до уваги можливість штучного втручання до і після народження.
Географічні розбіжності
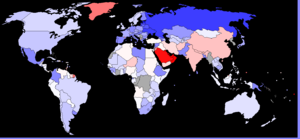
У таблиці наведено показники кількості чоловіків на 1 жінку в різних країнах..
| Держава/регіон | Новонароджені | 0-14 років | 15-24 роки | 25-54 роки | 55-64 роки | 65 і більше | Усе населення |
|---|---|---|---|---|---|---|---|
| Афганістан | 1.05 | 1.03 | 1.04 | 1.04 | 1.03 | 0.87 | 1.03 |
| Албанія | 1.11 | 1.12 | 1.05 | 0.91 | 0.98 | 0.89 | 0.98 |
| Алжир | 1.05 | 1.05 | 1.05 | 1.02 | 1.03 | 0.84 | 1.03 |
| Американське Самоа | 1.06 | 0.96 | 0.98 | 1.06 | 1.00 | 0.86 | 1.01 |
| Андорра | 1.07 | 1.05 | 1.08 | 1.06 | 1.07 | 1.00 | 1.07 |
| Ангола | 1.05 | 1.04 | 1.04 | 1.02 | 1.02 | 0.86 | 1.02 |
| Ангуїлла | 1.03 | 1.05 | 0.98 | 0.82 | 0.92 | 0.96 | 0.93 |
| Антигуа і Барбадос | 1.05 | 1.03 | 0.98 | 0.84 | 0.90 | 0.76 | 0.90 |
| Аргентина | 1.05 | 1.05 | 1.04 | 1.00 | 0.97 | 0.70 | 0.97 |
| Вірменія | 1.14 | 1.15 | 1.03 | 0.92 | 0.93 | 0.59 | 0.89 |
| Аруба | 1.02 | 1.01 | 1.01 | 0.93 | 0.90 | 0.65 | 0.90 |
| Австралія | 1.06 | 1.05 | 1.05 | 1.03 | 1.01 | 0.85 | 1.01 |
| Австрія | 1.05 | 1.05 | 1.04 | 1.01 | 0.95 | 0.73 | 0.95 |
| Азербайджан | 1.12 | 1.16 | 1.07 | 0.95 | 0.98 | 0.62 | 0.98 |
| Багами | 1.03 | 1.03 | 1.03 | 1.00 | 0.96 | 0.62 | 0.96 |
| Бахрейн | 1.03 | 1.03 | 1.28 | 1.90 | 1.54 | 0.91 | 1.54 |
| Бангладеш | 1.04 | 1.03 | 0.88 | 0.90 | 0.95 | 0.96 | 0.95 |
| Барбадос | 1.01 | 1.00 | 1.00 | 0.99 | 0.94 | 0.65 | 0.94 |
| Білорусь | 1.06 | 1.06 | 1.06 | 0.96 | 0.87 | 0.46 | 0.87 |
| Бельгія | 1.05 | 1.04 | 1.04 | 1.02 | 0.96 | 0.72 | 0.96 |
| Беліз | 1.05 | 1.04 | 1.04 | 1.02 | 1.03 | 0.90 | 1.03 |
| Бенін | 1.05 | 1.04 | 1.04 | 1.02 | 1.01 | 0.67 | 1.01 |
| Бермуди | 1.02 | 1.02 | 1.00 | 1.00 | 0.94 | 0.72 | 0.94 |
| Бутан | 1.05 | 1.04 | 1.04 | 1.15 | 1.09 | 1.11 | 1.10 |
| Болівія | 1.05 | 1.04 | 1.03 | 0.95 | 0.98 | 0.79 | 0.98 |
| Боснія і Герцоговина | 1.07 | 1.07 | 1.07 | 1.01 | 0.95 | 0.63 | 0.95 |
| Ботсвана | 1.03 | 1.04 | 0.99 | 1.12 | 1.03 | 0.67 | 1.02 |
| Бразилія | 1.05 | 1.04 | 1.03 | 0.98 | 0.97 | 0.74 | 0.98 |
| Британські Віргінські Острови | 1.05 | 0.97 | 0.91 | 0.90 | 0.92 | 0.95 | 0.93 |
| Бруней | 1.05 | 1.07 | 0.98 | 0.94 | 0.99 | 0.95 | 1.00 |
| Болгарія | 1.06 | 1.05 | 1.06 | 0.99 | 0.92 | 0.68 | 0.92 |
| Буркіна Фасо | 1.03 | 1.00 | 1.01 | 1.03 | 0.99 | 0.62 | 0.99 |
| Бірма | 1.06 | 1.04 | 1.03 | 0.99 | 0.99 | 0.77 | 0.99 |
| Бурунді | 1.03 | 1.01 | 1.00 | 1.00 | 0.99 | 0.67 | 0.98 |
| Кабо Верде | 1.03 | 1.01 | 1.00 | 0.94 | 0.94 | 0.60 | 0.94 |
| Камбоджа | 1.05 | 1.02 | 0.98 | 0.95 | 0.94 | 0.60 | 0.94 |
| Камерун | 1.03 | 1.02 | 1.01 | 1.01 | 1.01 | 0.84 | 1.01 |
| Канада | 1.06 | 1.05 | 1.06 | 1.03 | 0.99 | 0.79 | 0.99 |
| Кайманові Острови | 1.02 | 1.01 | 0.98 | 0.95 | 0.95 | 0.90 | 0.95 |
| Центрально-Африканська Республіка | 1.03 | 1.01 | 1.01 | 1.00 | 0.98 | 0.66 | 0.98 |
| Чад | 1.04 | 1.03 | 0.94 | 0.83 | 0.93 | 0.72 | 0.93 |
| Чилі | 1.04 | 1.04 | 1.04 | 0.99 | 0.97 | 0.71 | 0.97 |
| Китай | 1.11 | 1.16 | 1.13 | 1.05 | 1.06 | 0.92 | 1.06 |
| Колумбія | 1.06 | 1.05 | 1.04 | 0.98 | 0.98 | 0.72 | 0.98 |
| Комори | 1.03 | 0.99 | 0.94 | 0.90 | 0.94 | 0.92 | 0.94 |
| Конго | 1.03 | 1.02 | 1.01 | 1.00 | 0.99 | 0.72 | 0.99 |
| Республіка Конго | 1.03 | 1.02 | 0.99 | 1.03 | 1.01 | 0.69 | 0.99 |
| Острови Кука | 1.04 | 1.13 | 1.15 | 1.01 | 1.07 | 0.96 | 1.07 |
| Коста-Рика | 1.05 | 1.05 | 1.04 | 1.01 | 1.01 | 0.86 | 1.01 |
| Кот д'Івуар | 1.03 | 1.02 | 1.02 | 1.05 | 1.02 | 0.96 | 1.03 |
| Хорватія | 1.06 | 1.06 | 1.05 | 0.98 | 0.93 | 0.66 | 0.93 |
| Куба | 1.06 | 1.06 | 1.05 | 1.01 | 0.99 | 0.82 | 0.99 |
| Кюросао | 1.15 | 1.05 | 1.08 | 0.92 | 0.79 | 0.71 | 0.92 |
| Кіпр | 1.05 | 1.06 | 1.19 | 1.10 | 1.04 | 0.77 | 1.04 |
| Чехія | 1.06 | 1.06 | 1.05 | 1.05 | 0.97 | 0.67 | 0.95 |
| Данія | 1.06 | 1.05 | 1.04 | 1.00 | 0.97 | 0.80 | 0.97 |
| Джибуті | 1.03 | 1.01 | 0.89 | 0.71 | 0.85 | 0.82 | 0.86 |
| Домініка | 1.05 | 1.05 | 1.06 | 1.03 | 1.02 | 0.77 | 1.02 |
| Домініканська Республіка | 1.04 | 1.04 | 1.04 | 1.05 | 1.03 | 0.86 | 1.03 |
| Еквадор | 1.05 | 1.04 | 1.03 | 0.95 | 0.99 | 0.92 | 0.99 |
| Єгипет | 1.05 | 1.05 | 1.05 | 1.03 | 1.02 | 0.82 | 1.03 |
| Ель Сальвадор | 1.05 | 1.05 | 1.01 | 0.85 | 0.93 | 0.80 | 0.93 |
| Екваторіальна Гвінея | 1.03 | 1.03 | 1.04 | 0.98 | 0.99 | 0.75 | 0.99 |
| Еритрея | 1.03 | 1.01 | 1.00 | 0.98 | 0.98 | 0.79 | 0.98 |
| Естонія | 1.06 | 1.06 | 1.06 | 0.92 | 0.84 | 0.49 | 0.84 |
| Ефіопія | 1.03 | 1.00 | 0.99 | 0.99 | 0.99 | 0.83 | 0.99 |
| Європейський Союз | 1.06 | 1.05 | 1.05 | 1.02 | 0.94 | 0.74 | 0.96 |
| Фарерські острови | 1.07 | 1.08 | 1.07 | 1.18 | 1.09 | 0.93 | 1.09 |
| Фіджі | 1.05 | 1.05 | 1.05 | 1.05 | 1.03 | 0.85 | 1.03 |
| Фінляндія | 1.04 | 1.04 | 1.04 | 1.04 | 0.96 | 0.71 | 0.96 |
| Франція | 1.05 | 1.05 | 1.05 | 1.01 | 0.96 | 0.74 | 0.96 |
| Французька Полінезія | 1.05 | 1.06 | 1.07 | 1.05 | 1.05 | 0.95 | 1.05 |
| Габон | 1.03 | 1.01 | 1.00 | 1.00 | 0.99 | 0.73 | 0.99 |
| Гамбія | 1.03 | 1.01 | 0.98 | 0.96 | 0.98 | 0.90 | 0.98 |
| Газа Стріп | 1.06 | 1.06 | 1.05 | 1.05 | 1.04 | 0.68 | 1.04 |
| Грузія | 1.08 | 1.12 | 1.08 | 0.94 | 0.92 | 0.66 | 0.91 |
| Німеччина | 1.06 | 1.06 | 1.04 | 1.03 | 0.97 | 0.76 | 0.97 |
| Гана | 1.03 | 1.01 | 0.99 | 0.94 | 0.97 | 0.88 | 0.98 |
| Гібралтар | 1.07 | 1.05 | 1.09 | 1.01 | 1.01 | 0.95 | 1.01 |
| Греція | 1.06 | 1.06 | 1.04 | 0.99 | 0.96 | 0.78 | 0.96 |
| Гренландія | 1.05 | 1.03 | 1.03 | 1.12 | 1.10 | 1.10 | 1.11 |
| Гренада | 1.10 | 1.07 | 0.99 | 1.05 | 1.03 | 0.83 | 1.02 |
| Гуам | 1.06 | 1.06 | 1.07 | 1.04 | 1.03 | 0.84 | 1.03 |
| Гватемала | 1.05 | 1.04 | 1.01 | 0.91 | 0.97 | 0.87 | 0.97 |
| Гвінея | 1.03 | 1.02 | 1.02 | 1.01 | 1.00 | 0.79 | 1.00 |
| Гвінея-Бісау | 1.03 | 1.00 | 0.99 | 0.99 | 0.95 | 0.64 | 0.95 |
| Гаяна | 1.05 | 1.04 | 1.06 | 1.09 | 1.02 | 0.69 | 0.99 |
| Гаїті | 1.01 | 1.01 | 1.00 | 0.99 | 0.99 | 0.80 | 0.98 |
| Гондурас | 1.05 | 1.04 | 1.04 | 1.02 | 1.01 | 0.79 | 1.01 |
| Гонконг | 1.13 | 1.14 | 1.05 | 0.75 | 0.87 | 0.88 | 0.94 |
| Угорщина | 1.06 | 1.06 | 1.06 | 1.01 | 0.91 | 0.59 | 0.91 |
| Ісландія | 1.04 | 1.03 | 1.02 | 1.02 | 1.00 | 0.85 | 1.00 |
| Індія | 1.12 | 1.13 | 1.13 | 1.06 | 1.08 | 0.91 | 1.08 |
| Індонезія | 1.05 | 1.04 | 1.04 | 1.03 | 1.00 | 0.78 | 1.00 |
| Іран | 1.05 | 1.05 | 1.06 | 1.04 | 1.03 | 0.89 | 1.03 |
| Ірак | 1.05 | 1.04 | 1.03 | 1.04 | 1.03 | 0.87 | 1.03 |
| Ірландія | 1.06 | 1.04 | 1.04 | 1.01 | 1.00 | 0.84 | 1.00 |
| Острів Мен | 1.08 | 1.11 | 1.08 | 0.99 | 1.00 | 0.85 | 0.99 |
| Ізраїль | 1.05 | 1.05 | 1.05 | 1.05 | 1.01 | 0.78 | 1.01 |
| Італія | 1.06 | 1.05 | 1.01 | 0.98 | 0.93 | 0.74 | 0.93 |
| Ямайка | 1.05 | 1.04 | 1.01 | 0.97 | 0.98 | 0.81 | 0.98 |
| Японія | 1.06 | 1.07 | 1.09 | 0.98 | 0.94 | 0.76 | 0.95 |
| Джерсі | 1.06 | 1.07 | 1.04 | 1.00 | 0.97 | 0.74 | 0.96 |
| Йорданія | 1.06 | 1.05 | 1.05 | 1.01 | 1.02 | 0.95 | 1.03 |
| Казахстан | 0.94 | 1.00 | 1.04 | 0.95 | 0.92 | 0.52 | 0.92 |
| Кенія | 1.02 | 1.01 | 1.00 | 1.02 | 1.00 | 0.79 | 1.00 |
| Кірибаті | 1.05 | 1.04 | 1.00 | 0.93 | 0.96 | 0.64 | 0.96 |
| Корея, Північна | 1.05 | 1.03 | 1.02 | 1.00 | 0.94 | 0.51 | 0.94 |
| Корея, Південна | 1.07 | 1.08 | 1.13 | 1.04 | 1.00 | 0.69 | 1.00 |
| Косово | 1.08 | 1.08 | 1.11 | 1.12 | 1.06 | 0.73 | 1.06 |
| Кувейт | 1.05 | 1.08 | 1.22 | 1.74 | 1.42 | 0.96 | 1.43 |
| Киргизстан | 1.07 | 1.05 | 1.03 | 0.96 | 0.96 | 0.63 | 0.96 |
| Лаос | 1.04 | 1.02 | 0.99 | 0.97 | 0.99 | 0.82 | 0.99 |
| Латвія | 1.05 | 1.05 | 1.04 | 0.99 | 0.86 | 0.48 | 0.86 |
| Ліван | 1.05 | 1.05 | 1.04 | 1.02 | 1.00 | 0.86 | 0.96 |
| Лесото | 1.03 | 1.01 | 0.91 | 0.96 | 0.98 | 0.99 | 0.97 |
| Ліберія | 1.03 | 1.02 | 0.95 | 1.00 | 0.99 | 1.01 | 1.00 |
| Лівія | 1.05 | 1.05 | 1.07 | 1.10 | 1.08 | 1.04 | 1.08 |
| Ліхтенштейн | 1.26 | 1.14 | 0.98 | 1.00 | 0.99 | 0.81 | 0.98 |
| Литва | 1.06 | 1.05 | 1.05 | 1.00 | 0.89 | 0.53 | 0.89 |
| Люксембург | 1.07 | 1.06 | 1.05 | 1.00 | 0.97 | 0.71 | 0.97 |
| Макао | 1.05 | 1.10 | 1.08 | 0.81 | 0.91 | 0.88 | 0.91 |
| Македонія | 1.08 | 1.07 | 1.07 | 1.03 | 0.99 | 0.75 | 0.99 |
| Мадагаскар | 1.03 | 1.02 | 1.00 | 1.00 | 1.00 | 0.83 | 1.00 |
| Малаві | 1.02 | 0.99 | 0.99 | 0.98 | 0.98 | 0.75 | 0.99 |
| Малайзія | 1.07 | 1.06 | 1.03 | 1.03 | 1.03 | 0.89 | 1.03 |
| Мальдіви | 1.05 | 1.04 | 1.40 | 1.43 | 1.29 | 0.94 | 1.34 |
| Малі | 1.03 | 1.01 | 0.91 | 0.87 | 0.95 | 1.01 | 0.95 |
| Мальта | 1.06 | 1.05 | 1.06 | 1.04 | 0.99 | 0.79 | 0.99 |
| Маршалові Острови | 1.05 | 1.04 | 1.04 | 1.04 | 1.04 | 0.95 | 1.04 |
| Мавританія | 1.03 | 1.01 | 0.96 | 0.86 | 0.93 | 0.74 | 0.93 |
| Маврикій | 1.05 | 1.04 | 1.02 | 1.00 | 0.97 | 0.67 | 0.97 |
| Мексика | 1.05 | 1.05 | 1.02 | 0.93 | 0.96 | 0.81 | 0.96 |
| Мікронезія | 1.05 | 1.03 | 1.01 | 0.94 | 0.98 | 0.77 | 0.99 |
| Молдова | 1.06 | 1.07 | 1.07 | 1.00 | 0.95 | 0.60 | 0.94 |
| Монако | 1.04 | 1.05 | 1.05 | 0.99 | 0.95 | 0.81 | 0.95 |
| Монголія | 1.05 | 1.04 | 1.02 | 0.94 | 0.96 | 0.77 | 1.00 |
| Чорногорія | 1.07 | 0.95 | 0.89 | 1.17 | 0.99 | 0.66 | 0.99 |
| Марокко | 1.05 | 1.03 | 0.99 | 0.94 | 0.97 | 0.82 | 0.97 |
| Мозамбік | 1.02 | 1.01 | 0.95 | 0.88 | 0.95 | 0.85 | 0.95 |
| Намібія | 1.03 | 1.02 | 1.02 | 1.09 | 1.02 | 0.78 | 1.02 |
| Науру | 0.84 | 0.79 | 1.07 | 0.99 | 0.90 | 0.78 | 0.91 |
| Непал | 1.04 | 1.04 | 1.01 | 0.91 | 0.97 | 0.87 | 0.96 |
| Нідерланди | 1.05 | 1.05 | 1.04 | 1.01 | 0.98 | 0.80 | 0.98 |
| Нова Каледонія | 1.05 | 1.05 | 1.04 | 1.01 | 1.00 | 0.81 | 1.00 |
| Нова Зеландія | 1.05 | 1.05 | 1.05 | 1.00 | 0.99 | 0.85 | 0.99 |
| Нікарагуа | 1.05 | 1.04 | 1.01 | 0.90 | 0.96 | 0.83 | 0.96 |
| Нігер | 1.03 | 1.02 | 0.98 | 1.00 | 1.01 | 1.03 | 1.01 |
| Нігерія | 1.06 | 1.05 | 1.05 | 1.05 | 1.04 | 0.85 | 1.01 |
| Північні Марианські Острови | 1.06 | 1.09 | 1.23 | 0.73 | 0.93 | 0.91 | 0.93 |
| Норвегія | 1.06 | 1.05 | 1.06 | 1.06 | 1.01 | 0.78 | 0.98 |
| Оман | 1.05 | 1.05 | 1.10 | 1.41 | 1.20 | 1.03 | 1.22 |
| Пакистан | 1.05 | 1.06 | 1.07 | 1.08 | 1.06 | 0.89 | 1.06 |
| Палау | 1.05 | 1.07 | 0.99 | 1.56 | 1.10 | 0.40 | 1.12 |
| Панама | 1.05 | 1.04 | 1.04 | 1.03 | 1.01 | 0.86 | 1.02 |
| Папуа-Нова Гвінея | 1.05 | 1.04 | 1.03 | 1.07 | 1.05 | 1.14 | 1.05 |
| Парагвай | 1.05 | 1.04 | 1.01 | 1.00 | 1.01 | 0.87 | 1.01 |
| Перу | 1.05 | 1.04 | 1.00 | 0.93 | 0.97 | 0.90 | 0.97 |
| Філіппіни | 1.05 | 1.04 | 1.04 | 1.01 | 1.00 | 0.76 | 1.00 |
| Польща | 1.06 | 1.06 | 1.04 | 1.01 | 0.94 | 0.62 | 0.94 |
| Португалія | 1.07 | 1.09 | 1.13 | 1.02 | 0.95 | 0.69 | 0.95 |
| Пуерто-Рико | 1.02 | 1.05 | 1.04 | 0.91 | 0.92 | 0.77 | 0.92 |
| Катар | 1.02 | 1.03 | 2.75 | 4.80 | 3.37 | 1.50 | 3.29 |
| Румунія | 1.06 | 1.06 | 1.05 | 1.02 | 0.95 | 0.68 | 0.95 |
| Росія | 1.06 | 1.06 | 1.05 | 0.96 | 0.86 | 0.44 | 0.86 |
| Руанда | 1.03 | 1.02 | 1.00 | 1.01 | 0.99 | 0.67 | 0.99 |
| Самоа | 1.05 | 1.07 | 1.05 | 1.08 | 1.05 | 0.78 | 1.05 |
| Сан-Марино | 1.10 | 1.14 | 1.05 | 0.89 | 0.94 | 0.80 | 0.94 |
| Сан-Томе і Принсіпе | 1.03 | 1.04 | 1.03 | 0.96 | 1.00 | 0.84 | 1.00 |
| Саудівська Аравія | 1.05 | 1.05 | 1.15 | 1.33 | 1.20 | 1.08 | 1.21 |
| Сенегал | 1.03 | 1.01 | 0.99 | 0.84 | 0.94 | 0.85 | 0.94 |
| Сербія | 1.07 | 1.07 | 1.06 | 1.02 | 0.95 | 0.69 | 0.95 |
| Сейшели | 1.03 | 1.05 | 1.10 | 1.11 | 1.05 | 0.61 | 1.04 |
| С'єра-Леоне | 1.03 | 0.99 | 0.94 | 0.92 | 0.94 | 0.78 | 0.94 |
| Сінгапур | 1.07 | 1.05 | 0.97 | 0.95 | 0.96 | 0.82 | 0.96 |
| Словаччина | 1.07 | 1.05 | 1.06 | 1.02 | 0.94 | 0.60 | 0.94 |
| Словенія | 1.07 | 1.06 | 1.05 | 1.02 | 0.95 | 0.66 | 0.95 |
| Соломонові Острови | 1.05 | 1.06 | 1.06 | 1.04 | 1.04 | 0.95 | 1.04 |
| Сомалі | 1.03 | 1.00 | 1.03 | 1.07 | 1.01 | 0.66 | 1.01 |
| Південна Африка | 1.02 | 1.01 | 1.01 | 1.07 | 0.98 | 0.67 | 0.99 |
| Іспанія | 1.07 | 1.06 | 1.07 | 1.04 | 0.98 | 0.74 | 0.97 |
| Шрі Ланка | 1.04 | 1.04 | 1.03 | 0.96 | 0.96 | 0.75 | 0.96 |
| Судан | 1.05 | 1.03 | 1.07 | 0.94 | 1.02 | 1.24 | 1.02 |
| Суринам | 1.05 | 1.04 | 1.04 | 1.04 | 1.01 | 0.77 | 1.01 |
| Свазіленд | 1.03 | 1.02 | 1.02 | 1.06 | 1.00 | 0.69 | 0.99 |
| Швеція | 1.06 | 1.06 | 1.06 | 1.04 | 1.00 | 0.81 | 0.98 |
| Швейцарія | 1.06 | 1.06 | 1.04 | 1.01 | 0.97 | 0.76 | 0.97 |
| Сирія | 1.06 | 1.05 | 1.03 | 1.00 | 1.01 | 0.85 | 1.03 |
| Тайвань | 1.07 | 1.07 | 1.06 | 1.00 | 0.99 | 0.89 | 1.00 |
| Таджикистан | 1.05 | 1.04 | 1.03 | 0.98 | 0.99 | 0.73 | 0.99 |
| Танзанія | 1.03 | 1.02 | 1.00 | 1.01 | 0.99 | 0.76 | 0.99 |
| Таїланд | 1.05 | 1.05 | 1.04 | 0.97 | 0.97 | 0.82 | 0.98 |
| Того | 1.03 | 1.01 | 1.00 | 0.99 | 0.98 | 0.77 | 0.98 |
| Тринідад і Тобаго | 1.03 | 1.04 | 1.08 | 1.08 | 1.03 | 0.76 | 1.03 |
| Туніс | 1.07 | 1.07 | 1.01 | 0.95 | 0.99 | 0.96 | 0.99 |
| Туреччина | 1.05 | 1.05 | 1.04 | 1.02 | 1.02 | 0.84 | 1.02 |
| Туркменістан | 1.05 | 1.03 | 1.01 | 0.98 | 0.98 | 0.77 | 0.98 |
| Тувалу | 1.05 | 1.05 | 1.12 | 0.95 | 0.97 | 0.73 | 0.97 |
| Уганда | 1.03 | 0.99 | 0.99 | 1.00 | 0.99 | 0.80 | 0.99 |
| Україна | 1.07 | 1.06 | 1.04 | 0.94 | 0.85 | 0.49 | 0.85 |
| Об'єднані Арабські Емірати | 1.05 | 1.05 | 1.47 | 3.22 | 2.19 | 1.77 | 2.19 |
| Велика Британія | 1.05 | 1.05 | 1.04 | 1.04 | 0.99 | 0.80 | 0.99 |
| США | 1.05 | 1.05 | 1.05 | 1.00 | 0.97 | 0.77 | 0.97 |
| Уругвай | 1.04 | 1.04 | 1.03 | 0.97 | 0.93 | 0.66 | 0.93 |
| Узбекистан | 1.06 | 1.05 | 1.03 | 0.99 | 0.99 | 0.75 | 0.99 |
| Вануату | 1.05 | 1.04 | 0.99 | 0.96 | 1.00 | 1.07 | 1.01 |
| Венесуела | 1.05 | 1.04 | 1.01 | 0.97 | 0.98 | 0.79 | 0.98 |
| В'єтнам | 1.12 | 1.11 | 1.07 | 1.01 | 1.00 | 0.62 | 1.00 |
| Віргінські Острови | 1.06 | 1.02 | 0.85 | 0.83 | 0.87 | 0.83 | 0.88 |
| Західна Сахара | 1.04 | 1.02 | 1.01 | 0.97 | 0.99 | 0.78 | 0.98 |
| Ємен | 1.05 | 1.04 | 1.03 | 1.04 | 1.03 | 0.90 | 1.03 |
| Замбія | 1.03 | 1.01 | 1.00 | 1.02 | 1.00 | 0.75 | 1.00 |
| Зімбабве | 1.03 | 1.02 | 1.01 | 1.07 | 1.00 | 0.70 | 0.95 |
| Світ | 1.07 | 1.07 | 1.062 | 1.022 | 0.955 | 0.802 | 1.014 |
«Феномен воєнних років»
Значний дефіцит чоловічої статі з'являється під час і після воєн. У цей період в воюючих країнах спостерігається збільшення чоловічих народжень приблизно на 1-2%. Так, під час Першої світової війни в європейських країнах частка хлопчиків зросла на 1-2,5% порівняно зі звичайною і склала в Німеччині 108,5%, приблизно таке ж збільшення відбулося на кінець 1942 року у Великій Британії та Франції. В Закавказзі з початку 90-х років (конфлікт Вірменії і Азербайджану з приводу Нагірного Карабаху, грузино-абхазький конфлікт) відзначається зростання народжуваності хлопчиків. Коливання вторинного співвідношення статей в межах 116–119 характерні для всього Південно-Кавказького регіону Це явище, встановлене на дуже великому статистичному матеріалі, отримало в демографії назву «феномену воєнних років», оскільки співвідношення статей у людини в мирні роки досить стабільне.
Пониження третинного співвідношення статей в онтогенезі
В ході онтогенезу співвідношення статей у багатьох видів рослин, тварин і людини знижується. Це пов'язано з підвищеною смертністю і пошкоджуваністю чоловічих систем в порівнянні з відповідними жіночими. Ця картина спостерігається майже на всіх стадіях онтогенезу і на всіх рівнях організації, чи досліджуємо ми різні види (людина, тварини або рослини), різні рівні організації (особина, орган, тканину або клітину) або стійкість до різних шкідливих факторів середовища (низькі і високі температури, голод, отрута, паразити, хвороби та ін.).
Гамільтон (Hamilton, 1948) наводить огляд диференціальної смертності статей для 70 видів, включаючи такі різні форми життя, як нематоди, молюски, ракоподібні, комахи, павукоподібні, птахи, рептилії, риби і ссавці. Згідно з цими даними, у 62 видів (89%) середня тривалість життя самців коротша, ніж у самок; у більшості інших немає різниці, і лише в окремих випадках тривалість життя самців більша, ніж у самок.
Можна зробити висновок, що підвищена смертність чоловічої статі — загальнобіологічне явище, воно спостерігається у рослин, тварин і людини для всіх рівнів організації від усіх екстремальних значень факторів середовища.
У людини середня тривалість життя різна в різних країнах, і у чоловіків на 3-10 років менша, ніж у жінок.
Вплив генетичних факторів на вторинне співвідношення статей
Визначення статі дитини у людини не є чисто випадковою подією. Дослідженнями, проведеними в Саксонії в 1876–1885 рр. і пізніше в Англії, Франції, США і Фінляндії, було встановлено, що сім'ї, в яких превалює одна стать, з'являються значно частіше, а сім'ї, з рівним співвідношенням статей — набагато рідше в порівнянні з теоретичним очікуванням. Було також встановлено, що є слабка, але реальна тенденція в сім'ях набувати другу дитину тієї ж статі, що і перша, третіх дітей однакової статі з другими і т. д..
У великої рогатої худоби в потомстві окремих биків-виробників спостерігали переважання бичків (1.5%), а також кореляцію між співвідношенням статей в потомстві виробника і його батька.
Вплив репродуктивного рангу («успіху») на вторинне співвідношення статей
Репродуктивний ранг (доступ до шлюбних партнерів) батьків впливає на співвідношення статей їх потомства. У самців репродуктивний ранг корелює, як правило, з їх соціально-ієрархічним рангом. У самок може мати місце зворотна кореляція, оскільки їх ієрархічний ранг, як і у самців, визначається силою і агресивністю, а репродуктивний ранг — більше привабливістю і поступливістю. У оленів, свиней, овець, собак, тюленів і людей було показано, що самки, які мають, більший «репродуктивний успіх», народжували більше нащадків чоловічої статі.
Вплив факторів середовища на вторинне співвідношення статей
Кількість пилку і вторинне співвідношення статей у рослин
Було виявлено, що кількість пилку, що потрапляє на жіночу квітку, може впливати на вторинне співвідношення статей у перехрестнозапилюваних рослин. Цю залежність було підтверджено у чотирьох видів рослин, що належать до трьох родин — щавель (Rumex acetosa, Polygonaceae),дрімота (Melandrium album, Cariophyllaceae),конопля (Cannabis sativa, Cannabinaceae) і хміль (Humulus japonicus, Cannabinaceae).
Температура яєць
У багатьох видів черепах, ящірок, крокодилів і деяких змій стать потомства залежить від температури інкубації яєць. Так як самки визначають місце відкладання яєць, вони можуть контролювати стать потомства. Температура при якій досягається співвідношення статей 1: 1 називається пороговою температурою. У деяких видів черепах низькі температури інкубації призводять до появи самців, а високі — самок, у інших видів самці з'являються як при низьких, так і при високих температурах, а самки — при середніх.
Вторинне співвідношення статей та інтенсивність статевої діяльності
Інтенсивність (частота) статевої діяльності у тварин може впливати на вторинне співвідношення статей. Воно безпосередньо пов'язане з фізіологічними параметрами організму. Низька інтенсивність статевої діяльності у самців і самок рівнозначна тому, що в процесі запліднення беруть участь, як правило, більш старі спермії і яйцеклітини. У семи видів тварин, що належать до шести родин: (курей, мишей, кроликів, свиней, коней, баранів і великої рогатої худоби) при зменшенні статевої діяльності самців або старінні сперми спостерігалося зменшення числа народжень чоловічих особин.
Для людини збільшення народжуваності хлопчиків при підвищенні інтенсивності статевої діяльності чоловіків доведено на великому статистичному матеріалі в роботах Джеймса.
Відмінності в інактивації і загибелі Y- і X-сперміїв
Зв'язок вторинного співвідношення статей з інтенсивністю статевої діяльності у людини і різних видів тварин може бути зумовлений більш швидкою загибеллю або інактивацією Y-сперміїв у порівнянні з X-сперміями. Для людини це припущення було підтверджено експериментально. Було виявлено, що після тривалих періодів утримання вміст Y-хроматину в спермі помітно зменшується. При стриманості менше 2 днів відсоток Y-хроматину становив 43,5%, при утриманні 14 днів і більше — 37,2%. Зменшенням концентрації Y-сперматозоїдів у спермі з плином часу можна пояснити занижені у порівнянні з теоретично очікуваною величиною (50%) значення Y-хроматину, отримані різними авторами, а також великий розкид цих значень.
Старіння яйцеклітин
Багатьма авторами був відзначений зв'язок між затримкою запліднення яйцеклітин і вторинним співвідношенням статей. У класичних працях Ріхарда Гертвіга і Кушакевича на жабі (Rana esculenta) було відзначене підвищення народження чоловічих особин. Схожі результати були отримані на метеликах, шовкопряді, форелі, клопах, дрозофилі, мишах, щурах, кроликах, великій рогатій худобі і людях, всього 15 видів як з чоловічою, так і з жіночою гетерогаметністю, що належать до 11 родин.
Залежність вторинного співвідношення статей від віку матері
В. М. Большаков і Б. С. Кубанцев, проаналізувавши залежність співвідношення статей потомства від віку матері у лисиць, норок, песців, собак, свиней, овець, великої рогатої худоби, коней і людини дійшли висновку, що «… у молодих матерів в потомстві, як правило, переважають новонароджені чоловічої статі. У середньовікових матерів, що перебувають у розквіті своєї відтворювальної функції, зростає відносне число нащадків жіночої статі. У матерів старшої вікової групи знову збільшується в потомстві відсоток особин чоловічої статі.»
Залежність вторинного співвідношення статей від умов життя
Для цілих країн, чим кращі умови життя, добробут, клімат, харчування, тим менше народжується хлопчиків. Таку ж закономірність відзначають і тваринники — чим кращі умови утримання сільськогосподарських тварин, тим більше народжується самок.
Під час великих природних або соціальних зрушень (різкі зміни клімату, посуха, війна, голод, переселення) спостерігається тенденція підвищення вторинного співвідношення статей — росте відсоток хлопчиків.
Сучасна медицина дозволяє визначати стать пренатально. Це використовується в деяких країнах для вибору статі потомства, переважно шляхом абортування плодів небажаного статі, і може призводити навіть до порушення статево-вікової структури суспільства, чим пояснюється аномально велика кількість осіб чоловічої статі у віковій категорії до 30 років в сучасних Китаї і Індії (Див. також Біоетика).
Еволюційні механізми, що регулюють співвідношення статей в популяції
Початок робіт з еволюції співвідношення статей було покладено «принципом Фішера». Відповідно до цієї теорії, для видів, які вирощують своє потомство, третинне співвідношення статей 1 : 1 є оптимальним і забезпечує найкращі умови для виживання потомства. Щоб пояснити спостережувані у багатьох видів відхилення від цього співвідношення, Фішер стверджував, що рівним повинно бути не число самців і самок, а витрати батьків на відтворення потомства чоловічої або жіночої статі до завершення виховання потомства, тобто чим «дорожче обходяться» батькам нащадки даної статі, тим їх менше продукується. З «теорії рівних витрат» Фішера випливає, зокрема, що в тих випадках, коли нащадки різної статі мають різні розміри, повинно спостерігатися порушення співвідношення статей. Проте дані Хоува (Howe, 1977) по воронячих дроздах (Quiscalus quiscula), у яких самці важчі від самок на 20%, і велике дослідження Ньютона і Маркісса (Newton, Marquiss, 1978) по яструбу (Accipiter nisus), у якого, навпаки, самки вдвічі масивніше від самців, не підтвердили передбачення теорії Фішера. Гамільтон (Hamilton, 1967) звернув увагу на помилковість теорії Фішера у всіх випадках, коли має місце локальна конкуренція за схрещування. Він нарахував близько 25 видів кліщів і комах з 16 різних родин, у яких постійний значний інбридинг поєднується з великим надлишком самок і з арренотокічною системою розмноження.
На думку Калмусома і Сміта (Kalmus, Smith, 1960), значення теоретичного співвідношення статей 1 : 1 оптимальне, оскільки максимально полегшує зустріч особин протилежної статі і знижує ступінь інбридингу. Їх теорія не в змозі пояснити спостережувані відхилення значень вторинного співвідношення статей у багатьох видів від 1 : 1, а також його зміни в залежності від різних факторів.
Ближче від усіх до ідеї регуляції співвідношення статей підійшов Мейнард Сміт (1981), який припустив, що «батькам може бути вигідно продукувати потомство статі, більш рідкісної в даній місцевості».
Організменні механізми регуляції співвідношення статей
Сполучною ланкою зворотного зв'язку у перехрестнозапилюваних рослин служить кількість пилку, що потрапляє на жіночу квітку, а у тварин — інтенсивність статевої діяльності, яка проявляється через неоднакове старіння Х- і Y-сперміїв і через різну спорідненість до них свіжих і старих яйцеклітин. При цьому малі кількості пилку, інтенсивна статева діяльність самців, свіжа сперма і старі яйцеклітини — це ті чинники, які призводять до збільшення народжуваності чоловічих особин.
Популяційні механізми регуляції співвідношення статей
Для реалізації популяційного механізму необхідно, щоб ймовірність мати нащадка даної статі розрізнялася у різних особин і визначалася їх генотипом. При цьому повинна існувати зворотна залежність між репродуктивним рангом даної особини і статтю її потомства: чим вищий репродуктивний ранг, тим більше має бути нащадків протилежної статі. В цьому випадку регуляція може здійснюватися на популяційному рівні, більшою чи меншою участю в розмноженні особин, що дають в потомстві надлишок самців або самок.
Існування негативного зворотного зв'язку показано прямими експериментами принаймні у трьох видів (у одного виду рослин і двох видів тварин): Melandrium album, Lebistes reticulatus peters, Macrocheles. Багатьом видам рослин і тварин (4 види рослин і 16 видів тварин), а також людині притаманні ті чи інші механізми для його реалізації.
Див. також
|