
| Вірус | ||||||
|---|---|---|---|---|---|---|
 3-вимірна модель Ротавірусу
| ||||||
| Класифікація вірусів | ||||||
| ||||||
|
Групи
| ||||||
|
I: Дволанцюгові ДНК-віруси | ||||||
|
Посилання
| ||||||
|
Ві́рус (![]() ,лат. virus — отрута) — неклітинний інфекційний агент, який може відтворюватися лише всередині живих клітин. Віруси уражають всі типи організмів, від рослин і тварин до бактерій і архей (віруси бактерій зазвичай називають бактеріофагами). Виявлено також віруси, здатні реплікуватися лише за присутності інших вірусів (віруси-сателіти).
,лат. virus — отрута) — неклітинний інфекційний агент, який може відтворюватися лише всередині живих клітин. Віруси уражають всі типи організмів, від рослин і тварин до бактерій і архей (віруси бактерій зазвичай називають бактеріофагами). Виявлено також віруси, здатні реплікуватися лише за присутності інших вірусів (віруси-сателіти).
Від часу публікації 1892 року статті Дмитра Івановського, який описав небактеріальний патоген рослин тютюну, і відкриття 1898 року Мартіном Беєрінком вірусу тютюнової мозаїки було докладно описано понад 6 тисяч видів вірусів. Однак, припускають, що їх існує понад 100 мільйонів. Віруси виявлено майже в кожній екосистемі на Землі. Вони є найчисельнішою біологічною формою. Вивченням вірусів переймається наука вірусологія, розділ мікробіології.
У тварин вірусні інфекції спричинюють імунну відповідь, яка найчастіше призводить до знищення вірусу. Імунну відповідь також можна спричинити вакцинами, що дають активний набутий імунітет проти певної вірусної інфекції. Однак деяким вірусам, зокрема вірусу імунодефіциту людини і збудникам вірусних гепатитів, вдається проскочити повз імунну відповідь, спричинюючи хронічну хворобу. Антибіотики не діють на віруси, проте розроблено декілька противірусних препаратів.
Етимологія назви
Слово «вірус» утворене від лат. virus — «отрута». Аби позначити агент, здатний спричинити інфекційну хворобу, його вперше застосовано 1728 року, ще до відкриття вірусів, зробленого 1892 року Дмитром Івановським; він же ввів термін фільтрівний вірус як позначення небактеріального хвороботворного агента, здатного проходити крізь бактеріальні фільтри — фільтруватися. Термін «віріон», створення якого припадає на 1959 рік, застосовують щоби позначити одиничну стабільну вірусну частинку, яка залишила клітину й здатна інфікувати інші клітини такого ж типу.
Історія досліджень

Луї Пастер не зміг виявити мікроорганізм, що спричинює сказ, і припускав, що цей патоген занадто малий, щоб побачити його в мікроскоп. 1884 року французький мікробіолог Шарль Шамберлан винайшов фільтр (нині відомий як фільтр Шамберлана або фільтр Шамберлана—Пастера), шпарини якого менші за бактерії. За допомогою цього фільтра можна повністю видалити бактерії з розчину. 1892 року російський біолог Дмитро Івановський використовував його для вивчення виду, нині відомого як вірус тютюнової мозаїки. Його досліди показали, що екстракт перетертих листків заражених рослин тютюну зберігає інфекційні властивості й після фільтрації. Івановський припустив, що інфекцію може спричинити токсин, що виділяється бактеріями, однак він не розвинув цю думку. У той час вважали, що будь-який інфекційний агент можна виділити на фільтрі і вирощувати в живильному середовищі — це один з постулатів мікробної теорії хвороб. Крім того, Івановський в оптичному мікроскопі спостерігав у заражених клітинах рослин кристалоподібні тіла, які в сучасному розумінні були скупченнями вірусів, згодом їх названо «кристалами Івановського». 1898 року голландський мікробіолог Мартін Беєрінк повторив дослідження Івановського й зробив висновок, що інфекційний матеріал, який пройшов крізь фільтр, є не що інше, як нова форма інфекційних агентів. Він помітив, що агент розмножується тільки в клітинах котрі діляться, однак його досліди не виявили того, що саме являють собою частинки. Беєрінк назвав його розчинний живий мікроб (лат. Contagium vivum fluidum) і знову ввів у вжиток слово «вірус». Він твердив, що за своєю природою вірус рідкий. Згодом цю теорію спростував Венделл Стенлі, який довів, що віруси є частинками. Того ж року Фрідріх Лефлер і Пауль Фрош виявили перший вірус тварин — збудник ящуру (афтовірус), пропустивши його через схожий фільтр.
На початку XX століття англійський бактеріолог Фредерік Творт відкрив групу вірусів, котрі інфікують бактерії (нині вони відомі як бактеріофаги або просто фаги), а франко-канадський мікробіолог Фелікс Д'Ерелль описав віруси, які при додаванні до бактерій на агарі утворюють навколо себе простір з мертвими бактеріями. Д'Ерелль зробив точні розведення суспензії цих вірусів і встановив найбільше розведення (найменшу концентрацію вірусів), за якого не всі бактерії гинуть, але водночас утворюються окремі ділянки з мертвими клітинами. Підрахувавши кількість таких ділянок та враховуючи коефіцієнт розведення, він визначив кількість вірусних частинок у початковій суспензії. Фагів оголосили потенційним засобом від хвороб, таких як тиф і холера, проте увага до них зникла через відкриття більш ефективних пеніциліну та інших антибіотиків. Вивчення фагів дало розуміння про явище «вмикання» і «вимикання» генів, а також дозволило використовувати їх для запровадження в бактеріальний геном чужорідних генів.
Наприкінці XIX століття вже було відомо, що віруси здатні інфікувати, проходять крізь фільтри і потребують живого господаря для розмноження. Тоді віруси з дослідницькою метою культивували тільки в рослинах і тваринах. 1906 року Росс Ґренвілл Гаррісон винайшов спосіб вирощування тканин в лімфі, і 1913 року Штейнард, Ізраелі та Ламберт використовували цей метод, коли вирощували вірус осповакцини на фрагментах тканини рогівки морських свинок. 1928 року Р. Б. Мейтланд і М. К. Мейтланд виростили вірус осповакцини на суспензії з подрібнених курячих нирок. Цей метод не застосовували широко до кінця 1950-х років, коли у великих масштабах стали вирощувати поліовірус для виробництва вакцини.
Інше велике досягнення належить американському патологу Ернесту Вільяму Гудпасчеру: 1939 року він виростив вірус грипу та кілька інших вірусів у запліднених курячих яйцях. 1949 року Джон Франклін Ендерс, Томас Веллер і Фредерік Роббінс виростили поліовірус на культурі клітин зародка людини. Це був перший вірус, вирощений не на тканинах тварин або яйцях. Ця робота дала можливість Джонасу Солку створити дієву поліовакцину (вакцину проти поліомієліту).
Перші зображення вірусів отримали після того, як 1931 року німецькі інженери Ернст Руска і Макс Кнолль винайшли електронний мікроскоп. 1935 року американський біохімік і вірусолог Венделл Мередіт Стенлі ретельно вивчив вірус тютюнової мозаїки й виявив, що він здебільшого складається з білка. Невдовзі цей вірус було розділено на білкову та РНК-складову. Вірус тютюнової мозаїки було кристалізовано першим серед вірусів, що дозволило багато чого дізнатися про його структуру. Наприкінці 1930-х років Берналь і Фенкухен отримали першу рентгенограму кристалізованого вірусу. На підставі зроблених цим способом зображень Розалінд Франклін 1955 року визначила повну структуру цього вірусу. Того ж року Хейнц Людвіг Френкель-Конрат і Роблі Вільямс показали, що очищена РНК вірусу тютюнової мозаїки і білок його оболонки здатні до самоскладання у функціональний вірус. Це дозволило їм припустити, що подібний механізм лежить в основі складання вірусу всередині клітини-хазяїна.
Друга половина XX століття стала періодом розквіту вірусології. У той час було відкрито понад 2000 видів вірусів тварин, рослин і бактерій. 1957 року відкрито кінський артерівірус і збудник вірусної діареї корів (пестивірус). 1963 року Барух Бламберг відкрив вірус гепатиту B, а 1965 року Говард Темін описав перший ретровірус. 1970 року Темін і Девід Балтімор незалежно один від одного описали зворотну транскриптазу, ключовий фермент, за допомогою якого ретровіруси синтезують ДНК-копії своїх РНК. 1983 року група науковців на чолі з Люком Монтаньє з Інституту Пастера у Франції вперше виділила ретровірус, відомий нині як ВІЛ.
2002 року в Нью-Йоркському університеті створено перший синтетичний вірус — поліовірус.
Походження
|
| |
|---|---|
| 1. Звідки беруться нові віруси // Канал «Цікава наука» на YouTube, 30 січня 2021. | |
Віруси знайдено скрізь, де є життя, і, ймовірно, вони існують від миті появи перших живих клітин. Походження вірусів туманне, оскільки вони не залишають жодних викопних решток, а їхні родинні зв'язки можна вивчати тільки методами молекулярної філогенетики. Існує три основні гіпотези походження вірусів: регресивна гіпотеза, гіпотеза клітинного походження і гіпотеза коеволюції.
Регресивна гіпотеза
Згідно з цією гіпотезою, віруси колись були дрібними клітинами, що паразитують у більших клітинах. З плином часу ці клітини, ймовірно, втратили гени, які були «зайвими» за паразитичного способу життя. Це припущення ґрунтується на спостереженні, що деякі бактерії, а саме рикетсії та хламідії, є клітинними організмами, які, однак, подібно до вірусів, можуть розмножуватися тільки всередині іншої клітини. Цю гіпотезу також називають гіпотезою дегенерації або гіпотезою редукції.
Гіпотеза клітинного походження
Деякі віруси могли з'явитися з фрагментів ДНК або РНК, що «втекли» з геному більшого організму. Такі фрагменти можуть походити від плазмід (молекул ДНК, здатних передаватися від клітини до клітини) або від транспозонів (молекул ДНК, що реплікуються і пересуваються з місця на місце всередині геному). Транспозони, що їх раніше називали «генами, що стрибають», є прикладами мобільних елементів геному, можливо, від них могли піти деякі віруси. Їх відкрила Барбара Мак-Клінток 1950 року в кукурудзі. Цю гіпотезу також називають гіпотезою кочування або гіпотезою втечі.
Гіпотеза коеволюції
Згідно з цією гіпотезою, віруси походять від складних комплексів білків і нуклеїнових кислот одночасно з першими на Землі живими клітинами, і залежать від клітинного життя ось уже мільярди років. Крім вірусів, існують і інші неклітинні форми життя. Наприклад, віроїди — це молекули РНК, котрих не розглядають як віруси, через те що у них немає білкової оболонки. Утім, низка властивостей зближує їх з деякими вірусами, а тому їх відносять до субвірусних частинок. Віроїди є важливими патогенами рослин. Вони не кодують власні білки, однак взаємодіють із клітиною-хазяїном і використовують її, аби реплікувати свою РНК. Вірус гепатиту D має РНК-геном, подібний до геному віроїдів, проте сам не здатний синтезувати білок оболонки. Для утворення вірусних частинок він використовує білок капсиду вірусу гепатиту B і може розмножуватися лише в клітинах, заражених цим вірусом. Отже, вірус гепатиту D є дефектним вірусом. Подібно до цього вірофаг Супутник залежить від мімівірусу, який уражає найпростіше Acanthamoeba castellanii. Ці віруси залежать від наявності в клітині-хазяїні іншого вірусу і називаються вірусами-сателітами. Подібні віруси демонструють, як може виглядати проміжна ланка між вірусами і віроїдами.
У минулому кожна з цих гіпотез мала свої слабкі місця: регресивна гіпотеза не пояснювала, чому навіть найдрібніші клітинні паразити ніяк не схожі на віруси. Гіпотеза втечі не давала пояснення появи складних капсидів та інших складників вірусної частинки. Гіпотеза коеволюції суперечила визначенню вірусів як неклітинних частинок, залежних від клітин-хазяїв. Утім, нині багато фахівців визнають віруси стародавніми організмами, які з'явилися, імовірно, ще до поділу клітинного життя на три домени. Це відкриття дозволило сучасним вірусологам переглянути і переоцінити ці три класичні гіпотези.
Гіпотеза світу РНК та комп'ютерний аналіз послідовностей вірусної ДНК та ДНК хазяїна дають краще розуміння еволюційних взаємин між різними групами вірусів і можуть допомогти визначити предків сучасних вірусів. Допоки ці дослідження не прояснили, яка з трьох основних гіпотез правильна. Проте малоймовірно, щоби всі сучасні віруси мали спільного предка, і, можливо, в минулому віруси незалежно виникали кілька разів за одним або кількома механізмами, бо між різними групами вірусів є значні відмінності в організації генетичного матеріалу.

Пріони — це інфекційні білкові молекули, які не містять ДНК або РНК. Вони спричинюють такі захворювання, як скрепі в овечок, губчастоподібна енцефалопатія великої рогатої худоби та хронічна слабкість (англ. chronic wasting disease) у оленевих. До пріонових хвороб людини відносять куру, хворобу Кройтцфельда — Якоба та синдром Герстмана — Штройслера — Шейнкера. Пріони здатні заохочувати утворення власних копій. Пріоновий білок здатен існувати у двох ізоформах: нормальній (PrPC) та пріоновій (PrPSc). Пріонова форма, взаємодіючи з нормальним білком, сприяє його перетворенню на пріонову форму. Хоча пріони ґрунтовно відрізняються від вірусів і віроїдів, їх відкриття дає більше підстав повірити в те, що віруси могли піти від самовідтворюваних молекул.
Біологія
Віруси як форма життя
Поки вірус перебуває в позаклітинному середовищі або в процесі зараження клітини, він існує у вигляді незалежної частинки. Вірусні частинки (віріони) складаються з двох чи трьох компонентів: генетичного матеріалу у вигляді ДНК або РНК (деякі, наприклад мімівіруси, мають обидва типи молекул); білкової оболонки (капсиду), що захищає ці молекули, і, в деяких випадках, — додаткових ліпідних оболонок. Наявність капсиду відрізняє віруси від вірусоподібних інфекційних нуклеїнових кислот — віроїдів. Залежно від того, яким типом нуклеїнової кислоти представлений генетичний матеріал, виділяють ДНК-вмісні віруси і РНК-вмісні віруси; на цьому принципі ґрунтується класифікація вірусів за Балтімором. Раніше до вірусів також помилково відносили пріони, однак згодом виявилося, що ці збудники є особливими інфекційними білками і не містять нуклеїнових кислот. Форма вірусів різниться від простої спіральної та ікосаедричної до складніших структур. Розміри середнього вірусу становлять близько однієї сотої розміру середньої бактерії. Більшість вірусів занадто малі, щоби бути чітко помітними під оптичним мікроскопом.
Віруси є облігатними паразитами, тому що не здатні розмножуватися поза клітиною. Поза клітиною вірусні частинки не виявляють ознак живого і поводяться як частинки біополімерів. Від живих паразитарних організмів віруси відрізняються повною відсутністю основного енергетичного обміну і відсутністю найскладнішого елемента живих систем — апарату трансляції (синтезу білка), складність якого перевищує складність самих вірусів.
За одним із визначень віруси є формою життя, за іншим віруси є комплексами органічних молекул, що взаємодіють з живими організмами. Віруси визначають як «організми на межі живого». Віруси схожі на живі організми тим, що вони мають свій набір генів і еволюціюють шляхом природного добору, а також тим, що здатні розмножуватися, створюючи власні копії шляхом самоскладання. Віруси мають генетичний матеріал, однак позбавлені клітинної будови, а саме цю рису зазвичай розглядають як основоположну властивість живої матерії. У вірусів немає власного обміну речовин, і для синтезу власних молекул вони потребують клітини-хазяїна. Через це вони не здатні розмножуватися поза клітиною. Хоча такі бактерії, як рикетсії та хламідії, попри те, що вони не можуть розмножуватися поза клітиною хазяїна, вважають живими організмами. Загальновизнані форми життя розмножуються поділом клітини, тоді як вірусні частинки мимовільно збираються в інфікованій клітині. Від росту кристалів розмноження вірусів відрізняється тим, що віруси успадковують мутації і перебувають під тиском природного добору. Самоскладання вірусних частинок у клітині дає додаткове підтвердження гіпотези, що життя могло зародитися у вигляді органічних молекул, що самоскладаються. Опубліковані 2013 року дані про те, що деякі бактеріофаги мають власну імунну систему, здатну до адаптації, є додатковим доводом на користь визначення вірусу як форми життя.
Структура

Будова ікосаедричних віріонів:
А. Вірус, що не має ліпідної оболонки (наприклад, пікорнавірус). B. Оболонковий вірус (наприклад, герпесвірус). Цифрами позначено: (1) капсид, (2) геномна нуклеїнова кислота, (3) капсомер, (4) нуклеокапсид, (5) віріон, (6) ліпідна оболонка, (7) мембранні білки оболонки 
Палочкоподібний віріон вірусу тютюнової мозаїки. Цифрами позначено: (1) РНК-геном вірусу, (2) капсомер, що складається лише з одного протомера, (3) зріла ділянка капсиду
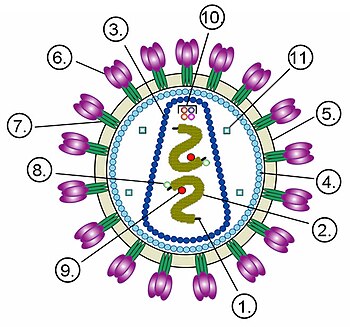
Будова віріона неікосаедричного оболонкового вірусу на прикладі ВІЛ. Цифрами позначено: (1) РНК-геном вірусу, (2) нуклеокапсид, (3) капсид, (4) білковий матрикс, що підстилає (5) ліпідну мембрану, (6) gp120 — глікопротеїн, за допомогою якого відбувається зв'язування вірусу з клітинною мембраною, (7) gp41 — глікопротеїн.
Цифрами 8-11 позначено білки, що входять до складу віріона й потрібні вірусу на ранніх стадіях інфекції: (8) — інтеграза, (9) — зворотна транскриптаза, (10) — Vif, Vpr, Nef і p7, (11) — протеаза |
Віруси дуже різняться за формою та розміром. Зазвичай, вони значно дрібніші за бактерії. Більшість вивчених вірусів мають діаметр у межах від 20 до 300 нм. Деякі філовіруси завдовжки до 1400 нм, а їхній діаметр становить лише 80 нм. 2013 року найбільшим з відомих вірусів вважали пандоравірус розмірами 1 × 0,5 мкм, однак 2014 року з багаторічної мерзлоти з Сибіру описано Pithovirus, що сягає 1,5 мкм завдовжки і 0,5 мкм у діаметрі. Нині його вважають найбільшим з відомих вірусів. Більшість віріонів неможливо побачити в оптичний мікроскоп, тому використовують електронні — і сканувальні, і трансмісійні. Аби віруси різко виділялися на навколишньому тлі, застосовують електроннощільні «барвники». Вони є розчинами солей важких металів, наприклад, вольфраму, що розсіюють електрони на покритій ними поверхні. Однак обробка такими речовинами погіршує видимість дрібних деталей. Цю проблему вирішує негативне контрастування, коли «забарвлюють» лише тло.
Окрема вірусна частинка, відома як віріон, складається з нуклеїнової кислоти, покритої захисною білковою оболонкою — капсидом. Капсид складається з однакових білкових субодиниць, які називаються капсомерами. Капсид, як правило, є ліпофільним, тобто має високу спорідненість до клітинних ліпідних мембран, цим полегшується проникнення вірусу до клітини. Також поверхня вірусів може містити «вирости», зазвичай глікопротеїни, які розпізнають особливі для них рецептори на поверхні клітини-мішені[джерело?].
Віруси можуть також мати ліпідну оболонку навколо капсиду (суперкапсид), утворену з мембрани клітини-хазяїна. Капсид складається з білків, що кодуються вірусним геномом, а його форма лежить в основі класифікації вірусів за морфологічною ознакою. Складноорганізовані віруси, крім того, кодують спеціальні білки, які допомагають у складанні капсиду. Комплекси білків і нуклеїнових кислот відомі як нуклеопротеїни, а комплекс білків вірусного капсиду з вірусною нуклеїновою кислотою називається нуклеокапсидом. Форму капсиду і віріона в цілому можна механічно (фізично) дослідити за допомогою атомно-силового мікроскопа. Самі ж поверхневі білки є антигенами, тобто розпізнаються клітинами імунної системи організму і дають сигнал для вироблення специфічних антитіл. Афінність антитіл визначається міцністю їх зв'язування із антигеном за рахунок сукупності всіх міжмолекулярних сил відштовхування та тяжіння[джерело?].
Капсид
Класифікують чотири морфологічні типи капсидів вірусів: спіральний, ікосаедричний, довгастий і комплексний.
Спіральний
Ці капсиди складаються з одного типу капсомерів, укладених спіраллю навколо центральної осі. У центрі цієї структури може бути порожнина або канал. Така організація капсомерів призводить до утворення паличкоподібних і ниткоподібних віріонів: вони можуть бути короткими і дуже жорсткими або довгими і дуже гнучкими. Генетичний матеріал, зазвичай представлений одноланцюговою РНК (часом одноланцюговою ДНК), утримується в білковій спіралі іонними взаємодіями між негативними зарядами на нуклеїнових кислотах і позитивними зарядами на білках. Загалом, довжина спірального капсиду залежить від довжини оточеної ним нуклеїнової кислоти, а діаметр визначається розміром і розташуванням капсомерів. Прикладом спірального вірусу є вірус тютюнової мозаїки.

Ікосаедричний
Більшість вірусів тварин мають ікосаедричну або майже кулясту форму з ікосаедричною симетрією. Правильний ікосаедр є оптимальною формою для закритого капсиду, складеного з однакових субодиниць. Мінімально необхідна кількість однакових капсомерів — 12, кожен капсомер складається з п'яти ідентичних субодиниць. Багато вірусів, як-от ротавірус, мають понад дванадцять капсомерів і виглядають круглими, але зберігають ікосаедричну симетрію. Капсомери, що лежать у вершинах, оточені п'ятьма іншими капсомерами і називаються пентонами. Капсомери трикутних граней мають 6 сусідів-капсомерів і називаються гексонами. Гексони, по суті, є плоскими, а пентони, що утворюють 12 вершин, — ламаними. Один і той самий білок може бути субодиницею і пентомерів, і гексамерів, або ж вони можуть складатися з різних білків.
Довгастий
Довгастими називають ікосаедричні капсиди, витягнуті вздовж осі симетрії п'ятого порядку. Така форма характерна для головок бактеріофагів.
Комплексний
Форма цих капсидів ні суто спіральна, ні суто ікосаедрична. Вони можуть нести додаткові зовнішні структури, приміром, білкові хвости або складні зовнішні стінки. Деякі бактеріофаги, як-от фаг Т4, мають комплексний капсид, що складається з ікосаедричної головки, з'єднаної зі спіральним хвостом, який може мати шестигранну основу з хвостовими білковими нитками, що відходять від неї. Цей хвіст діє на зразок молекулярного шприца, прикріплюючись до клітини-господаря і потім впорскуючи в неї генетичний матеріал вірусу.
Суперкапсид
Деякі віруси оточують себе додатковою оболонкою з модифікованої клітинної мембрани (плазматичної або внутрішньої, як-от ядерна мембрана або мембрана ендоплазматичного ретикулуму). Цей додатковий біліпідний шар називається суперкапсидом. У ліпідній оболонці вірусу також знаходяться білки, закодовані в вірусному геномі та/чи геномі хазяїна. Сама ж мембрана, а також будь-які її вуглеводні компоненти походять винятково від клітини-хазяїна. Таким чином формують свою оболонку вірус грипу і ВІЛ. Інфекційність більшості вірусів, що мають оболонку, залежить саме від цієї оболонки.
До вірусів з суперкапсидом належать поксвіруси, мімівіруси, мегавірус тощо. Поксвіруси — це великі складноорганізовані віруси з незвичайною морфологією. Всередині віріона знаходиться зв'язана з білками дволанцюгова молекула ДНК у вигляді дископодібної структури — нуклеоїда. Нуклеоїд оточений ліпідною мембраною та двома боковими тільцями невідомої функції. Вірус має суперкапсид з великою кількістю білків на його поверхні. Весь віріон злегка плейоморфний (тобто здатний змінювати форму і розмір залежно від умов) і може набувати форми від овальної до блокоподібної. Мімівірус є одним з найбільших описаних вірусів і має ікосаедричний капсид діаметром 400—500 нм. Білкові філаменти, що відходять від поверхні віріона, сягають 100 нм завдовжки. 2011 року дослідники виявили ще більший вірус на океанічному дні поблизу узбережжя Чилі. Вірус, якому дали тимчасову назву Megavirus chilensis, можна побачити навіть у звичайний оптичний мікроскоп.
Геном
| Властивості | Параметри |
|---|---|
| Нуклеїнова кислота |
|
| Форма |
|
| Кількість ланцюгів |
|
| Полярність |
|

Віруси показують величезну кількість варіантів організації геному; в цьому сенсі вони різноманітніші, ніж рослини, тварини, археї та бактерії. Існують мільйони різних типів вірусів, але тільки приблизно 5000 з них детально описані. Генетичний матеріал вірусу може бути представлений або ДНК, або РНК, відповідно, віруси поділяють на ДНК-вмісні і РНК-вмісні. Переважна більшість вірусів є РНК-вмісними. Віруси рослин найчастіше містять одноланцюгову РНК, а бактеріофаги, як правило, мають дволанцюгові ДНК.
Вірусний геном буває кільцевим, як у поліомавірусів, або лінійним, як у аденовірусів. Форма геному не залежить від типу нуклеїнової кислоти. У багатьох РНК-вмісних вірусів і деяких ДНК-вмісних вірусів геном часто представлений декількома окремими молекулами в кожному віріоні, у зв'язку з чим його називають сегментованим. У РНК-вмісних вірусів кожен сегмент (молекула) часто кодує тільки один білок, і зазвичай ці сегменти упаковуються в один капсид. Проте наявність усіх сегментів не завжди обов'язкова для інфекційності вірусу, як це демонструють вірус мозаїки стоколоса та деякі інші віруси рослин.
Вірусні геноми незалежно від типу нуклеїнової кислоти майже завжди бувають або одноланцюговими, або дволанцюговими. Дволанцюговий геном включає пару комплементарних ланцюгів нуклеїнової кислоти, а одноланцюговий — лише один ланцюг. Геном вірусів деяких родин (наприклад, гепаднавіруси) частково одноланцюговий і частково дволанцюговий.
Для більшості РНК-вмісних вірусів і деяких вірусів з одноланцюгової ДНК визначають полярність нуклеїнової кислоти залежно від того, чи комплементарна вона вірусній мРНК. Молекула РНК з позитивною полярністю (плюс-ланцюг) має ту ж послідовність нуклеотидів, що й мРНК, через те, принаймні, якась її частина може одразу ж почати транслюватися клітиною-господарем. РНК з негативною полярністю (мінус-ланцюг) комплементарна мРНК, тому до початку трансляції на ній має бути синтезована позитивна РНК за допомогою ферменту РНК-залежної-РНК-полімерази. Назви ланцюгів ДНК для вірусів, що містять одноланцюгову ДНК, подібні до назв ланцюгів для РНК: ланцюг, що кодує, комплементарний мРНК (-), а той, що не кодує, є її копією (+). Однак геноми декількох типів ДНК — і РНК-вмісних вірусів представлені молекулами, що мають різну полярність, тобто транскрипції може підлягати будь-який ланцюг. Це стосується, наприклад, гемінівірусів — вірусів рослин, що містять одноланцюгову ДНК, — і аренавірусів — вірусів тварин з одноланцюговою РНК.
Розмір геному широко різниться в різних видів. Найменший одноланцюговий ДНК-геном має вірус з родини цирковірусів: його геном кодує лише два білки і містить лише 2000 нуклеотидів. Один з найбільших геномів виявлено в мімівірусу: він містить понад 1,2 млн пар основ і кодує понад тисячу білків. Зазвичай, РНК-вмісні віруси мають менший геном, ніж ДНК-вмісні — розмір їхнього геному обмежений через більшу ймовірність помилок під час реплікації. За більшого розміру геному помилки, що відбулися під час його реплікації, зробили б вірус нежиттєздатним чи неконкурентоспроможним. Щоби подолати це обмеження, РНК-віруси часто мають сегментований геном — це зменшує ймовірність того, що помилка в одному з сегментів стане фатальною для всього геному. Натомість, ДНК-вмісні віруси зазвичай мають більші геноми завдяки більшій точності їх реплікативних ферментів. Однак віруси, що містять одноланцюгові ДНК, є винятком з цього правила — швидкість накопичення мутацій в їхніх геномах наближається до швидкості для вірусів, що містять одноланцюгові РНК.
Генетичні зміни відбуваються у вірусів за різними механізмами. До них належать випадкові заміни окремих основ у РНК або ДНК. У більшості випадків ці точкові мутації є «такими, що мовчать» — вони не змінюють структуру білків, що кодуються мутантними генами, але іноді внаслідок таких змін вірус може набути еволюційних переваг, як-от стійкість до противірусних препаратів. Дрейф антигенів відбувається тоді, коли геном вірусу зазнає значних змін. Це може бути наслідком рекомбінації або реасортименту. Коли це трапляється з вірусом грипу, наслідком може стати пандемія. РНК-віруси часто існують як квазівиди або суміш вірусів одного виду, але з трохи різними нуклеотидними послідовностями геному. Такі квазівиди є головною мішенню для природного добору.
Сегментований геном дає еволюційні переваги: різні штами вірусу з сегментованим геномом можуть обмінюватися генами і виробляти нащадків з унікальними властивостями. Це явище називається реасортиментом.
Генетична рекомбінація — це процес внесення розриву в молекулу нуклеїнової кислоти з наступним «зшиванням» її з іншими молекулами нуклеїнової кислоти. Рекомбінація може відбуватися між геномами двох вірусів, коли вони заражають клітину одночасно. Дослідження еволюції вірусів показали, що у вивчених видів рекомбінація досить поширена. Рекомбінація характерна і для РНК- і для ДНК-вмісних вірусів.
Життєвий цикл

Віруси не розмножуються клітинним поділом, оскільки не мають клітинної будови. Замість цього вони використовують ресурси клітини-хазяїна для утворення множинних копій самих себе, і їх складання відбувається всередині клітини.
Умовно життєвий цикл вірусу можна розбити на кілька етапів, що взаємно перекриваються (зазвичай виділяють 6 етапів):
- Прикріплення — це утворення специфічного зв'язку між білками вірусного капсиду і рецепторами на поверхні клітини-хазяїна. Це особливе зв'язування визначає коло хазяїнів вірусу. Наприклад, ВІЛ вражає тільки певний тип людських лейкоцитів. Це пов'язано з тим, що оболонковий глікопротеїн вірусу gp120 специфічно зв'язується з молекулою CD4 — хемокіновим рецептором, який зазвичай зустрічається на поверхні CD4+ T-лімфоцитів. Цей механізм забезпечує інфікування вірусом тільки тих клітин, які здатні здійснити його реплікацію. Зв'язування з рецептором може спричинити конформаційні зміни білків оболонки (або білка капсиду в разі безоболонкового вірусу), що водночас є сигналом до злиття вірусної і клітинної мембран і проникнення вірусу в клітину.
- Проникнення в клітину. На наступному етапі вірус має доправити всередину клітини свій генетичний матеріал. Деякі віруси вносять також власні білки, необхідні для її реалізації (особливо це притаманно для вірусів, які містять негативні РНК). Різні віруси для проникнення в клітину використовують відмінні стратегії: наприклад, пікорнавіруси впорскують свою РНК крізь плазматичну мембрану, а віріони ортоміксовірусів захоплюються клітиною в ході ендоцитозу й потрапляють в кисле середовище лізосом, де відбувається депротеїнізація вірусної частинки, після чого РНК в сукупності з вірусними білками долає лізосомальну мембрану і проходить у цитоплазму. Віруси також розрізняють за тим, де відбувається їхня реплікація: частина вірусів (наприклад, ті ж пікорнавіруси) розмножується в цитоплазмі клітини, а частина (наприклад, ортоміксовіруси) в її ядрі. Хід інфікування вірусами клітин грибів і рослин відрізняється від інфікування клітин тварин. Рослини мають міцну клітинну стінку, що складається з целюлози, а гриби — з хітину, тож більшість вірусів можуть пройти в них лише після ушкодження клітинної стінки. Однак майже всі віруси рослин (зокрема вірус тютюнової мозаїки) можуть переміщатися з клітини в клітину у формі одноланцюгових нуклеопротеїнових комплексів через плазмодесми. Бактерії, як і рослини, мають міцну клітинну стінку, яку вірус, щоби потрапити всередину, має пошкодити. Але через те, що клітинна стінка бактерій набагато тонша, ніж у рослин, деякі віруси виробили механізм впорскування геному в бактеріальну клітину через перетин клітинної стінки, за якого капсид залишається зовні.
- Позбавлення оболонок — це процес втрати капсиду. Це досягається за допомогою вірусних ферментів або ферментів клітини-господаря, а може бути і наслідком простої дисоціації. Зрештою вірусна геномна нуклеїнова кислота вивільняється.
- Реплікація вірусів передбачає насамперед реплікацію геному. Реплікація вірусу запускає синтез мРНК ранніх генів вірусу (з винятками для вірусів, що містять позитивну РНК), синтез вірусних білків, можливо, складання складних білків і реплікацію вірусного геному, яка запускається після збудження ранніх або регуляторних генів. Після цього може йти (у комплексних вірусів з великими геномами) ще одне або кілька кіл додаткового синтезу мРНК: «пізня» експресія генів призводить до синтезу структурних або віріонних білків.

- Після цього відбувається складання вірусних частинок, згодом стаються деякі модифікації білків. У вірусів, таких як ВІЛ, такі перетворення (іноді звані дозріванням) відбуваються після виходу вірусу з клітини-господаря.
- Вихід із клітини. Віруси можуть покинути клітину після лізису, процесу, під час якого клітина гине через розрив мембрани і клітинної стінки, якщо вона є. Ця особливість є у багатьох бактеріальних і деяких тваринних вірусів. Деякі віруси підлягають лізогенному циклу, де вірусний геном залучається шляхом генетичної рекомбінації у спеціальне місце хромосоми клітини-господаря. Тоді вірусний геном називають провірусом, або, у разі бактеріофага, профагом. Коли клітина ділиться, вірусний геном також подвоюється. У межах клітини вірус переважно не виявляє себе; однак у певну мить провірус або профаг може спричинити активацію вірусу, який здатен спричинити лізис клітин-господарів.
Вірус, що активно розмножується, не завжди вбиває клітину-господаря. Оболонкові віруси, зокрема ВІЛ, зазвичай відокремлюються від клітини шляхом брунькування. У ході цього процесу вірус отримує свою оболонку, що є модифікованим фрагментом клітинної мембрани господаря або іншої внутрішньої мембрани. Отже, клітина може продовжувати жити і виробляти вірус.
Особливості життєвого циклу різних груп
Генетичний матеріал всередині вірусних частинок і спосіб його реплікації значно відрізняється у різних вірусів.
- ДНК-вмісні віруси. Реплікація геному в більшості ДНК-вмісних вірусів відбувається в клітинному ядрі. Якщо клітина має відповідний рецептор на своїй поверхні, ці віруси проходять у клітину або шляхом безпосереднього злиття з клітинною мембраною (напр. герпесвіруси), або, що буває частіше — шляхом рецептор-залежного ендоцитозу. Більшість ДНК-вмісних вірусів повністю покладаються на синтетичний апарат клітини-господаря для виробництва їхнього ДНК і РНК, а також подальшого процесингу РНК. Однак віруси з великими геномами (наприклад, поксвіруси) можуть самостійно кодувати більшу частину необхідних для цього білків. Геном вірусу еукаріотів має подолати ядерну оболонку для того, щоб отримати доступ до ферментів, які синтезують ДНК і РНК, у разі ж бактеріофагів йому досить просто пройти в клітину.
- РНК-вмісні віруси. Реплікація таких вірусів зазвичай відбувається в цитоплазмі. РНК-вмісні віруси можна поділити на 4 групи в залежності від способу їхньої реплікації. Механізм реплікації визначається тим, чи є геном вірусу одноланцюговим чи дволанцюговим, другим важливим чинником у разі одноланцюгового геному є його полярність (чи може він безпосередньо бути матрицею для синтезу білка рибосомами). Всі РНК-віруси використовують власну РНК-репліказу для копіювання своїх геномів.
- Віруси, що використовують зворотну транскрипцію. Ці віруси містять одноланцюгову РНК (ретровіруси, Metaviridae, Pseudoviridae) або дволанцюгову ДНК (каулімовіруси і гепаднавіруси). РНК-вмісні віруси, здатні до зворотної транскрипції (ретровіруси, наприклад, ВІЛ), використовують ДНК-копію геному як проміжну молекулу під час реплікації, а ті, що містять ДНК (параретровіруси, наприклад, вірус гепатиту B) — РНК. В обох випадках використовується зворотна транскриптаза, або РНК-залежна ДНК-полімераза. Ретровіруси вбудовують ДНК, що утворюється у процесі зворотної транскрипції, в геном господаря, такий стан вірусу називають провірусом. Параретровіруси ж цього не роблять, хоча вбудовані копії їхнього геному можуть давати початок інфекційним вірусам, особливо у рослин. Віруси, що використовують зворотну транскрипцію, сприйнятливі до противірусних препаратів, що інгібують зворотну транскриптазу, зокрема до зідовудину та ламівудину.
Дія на клітини

Діапазон структурних та біохімічних ефектів, що спричинює вірус в інфікованій клітині, дуже широкий. Їх називають цитопатичними ефектами. Більшість вірусних інфекцій призводять до загибелі клітин-хазяїв. Причинами загибелі можуть бути лізис клітини, зміни клітинної мембрани і апоптоз. Часто причиною загибелі клітини є придушення її нормальної активності білками вірусу, не всі з яких входять до складу вірусної частинки.
Деякі віруси не спричинюють жодних видимих змін в ураженій клітині. Клітини, в яких вірус перебуває в латентному стані і неактивний, мають мало ознак інфекції і нормально функціюють. Це є причиною хронічних інфекцій, і вірус за них може ніяк себе не проявляти багато місяців або років. Так часто буває, наприклад, з вірусом герпесу. Деякі віруси, наприклад вірус Епштейна — Барр, можуть спричинити швидке розмноження клітин без появи злоякісності, тоді як інші, як-от папіломавіруси, можуть породити рак.
Діапазон хазяїв
Віруси, безперечно, є найчисленнішими біологічними об'єктами на Землі, і за цим показником вони перевершують усі організми, разом узяті. Вони вражають всі форми клітинних організмів, включаючи тварин, рослини, бактерії та гриби. Утім, різні типи вірусів можуть вражати тільки обмежене коло хазяїв, багато вірусів видоспецифічні. Деякі, як, наприклад, вірус віспи, можуть вражати тільки один вид — людей, у таких випадках кажуть, що вірус має вузький діапазон хазяїв. Навпаки, вірус сказу може вражати різні види ссавців, тобто він має широкий діапазон хазяїв. Віруси рослин нешкідливі для тварин, а більшість вірусів тварин безпечні для людини. Діапазон господарів деяких бактеріофагів обмежується одним штамом бактерій, і їх можна використовувати для визначення штамів, що спричинюють спалахи інфекційних захворювань методом фагового типування.
Поширення
Віруси поширюються багатьма способами: віруси рослин часто передаються від рослини до рослини комахами, що харчуються рослинними соками, наприклад, попелицями; віруси тварин можуть поширюватися комахами, такі організми відомі як переносники. Вірус грипу поширюється повітряно-крапельним шляхом під час кашлю та чхання. Норовірус і ротавірус, що зазвичай спричинюють вірусні гастроентерити, передаються фекально-оральним шляхом під час контакту з зараженою їжею або водою. ВІЛ є одним з декількох вірусів, що передаються статевим шляхом і під час переливання зараженої крові. Кожен вірус має певну специфічність до хазяїв, що визначається типами клітин, які він може інфікувати. Коло господарів може бути вузьким або, якщо вірус вражає багато видів, широким.
Класифікація
У біологічній класифікації віруси виділяють в окремий таксон, який утворює в класифікації Systema Naturae 2000 разом з доменами Bacteria, Archaea і Eukaryota кореневий таксон Biota. Протягом XX століття в систематиці висували пропозиції про створення виділеного таксона для неклітинних форм життя (Aphanobionta Novak, 1930; надцарство Acytota Jeffrey, 1971; Acellularia), проте такі пропозиції не були кодифіковані.
Головним завданням класифікації є опис різних вірусів і групування їх на підставі спільних властивостей. 1962 року Андре Львов, Роберт Горн і Пауль Турньє були першими, хто розробив основні принципи класифікації вірусів на підставі Ліннеївської ієрархічної системи. Основними таксонами в цій системі є тип, клас, ряд, родина, рід і вид. Віруси було розділено на групи за загальними властивостями (але не властивостями їхніх хазяїв) і типом нуклеїнових кислот у геномах. Пізніше створено Міжнародний комітет з таксономії вірусів. Однак у таксономії вірусів не застосовують поняття «царство», «відділ» і «клас», оскільки їхній малий розмір геному і висока частота мутацій ускладнює з'ясування спорідненості груп старших за порядок. По суті, класифікація вірусів за Балтімором є доповненням традиційнішої класифікації.
Систематику та таксономію вірусів нині кодифікує і підтримує Міжнародний комітет з таксономії вірусів (International Committee on Taxonomy of Viruses, ICTV), що підтримує також і таксономічну базу The Universal Virus Database ICTVdB.
Класифікація ICTV
Міжнародний комітет з таксономії вірусів розробив сучасну класифікацію вірусів і виділив основні властивості вірусів, що мають більшу вагу для класифікації зі збереженням одноманітності родин.
Розроблено об'єднану таксономію (універсальну систему для класифікації вірусів). Сьомий звіт ICTV закріпив для початку поняття про вид вірусу як про найнижчий таксон в ієрархії вірусів. Однак дотепер вивчено лише невелику частину від загального різноманіття вірусів, аналіз зразків вірусів з людського організму виявив, що близько 20 % послідовностей вірусних нуклеїнових кислот ще не було розглянуто раніше, а зразки з навколишнього середовища, наприклад, морської води і океанського дна, показали, що переважна більшість послідовностей є абсолютно новими.
Основними таксономічними одиницями є:
Сучасна класифікація ICTV (2017 року) містить 9 порядків вірусів: Bunyavirales, Caudovirales, Herpesvirales, Ligamenvirales, Mononegavirales, Nidovirales, Ortervirales, Picornavirales і Tymovirales. Існування ще одного порядку (Megavirales) тільки припускають. Класифікація не виділяє підвиди, штами та ізоляти. Загалом налічується 9 порядків, 127 родин, 44 підродини, 782 роди, 4686 видів і понад 3000 ще не класифікованих вірусів.
Класифікація за Балтімором
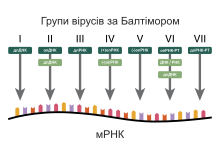
Лауреат Нобелівської премії біолог Девід Балтімор розробив класифікацію вірусів за Балтімором. Класифікація ICTV нині поєднується з класифікацією за Балтімором, становлячи сучасну систему класифікації вірусів.
Класифікація вірусів за Балтімором ґрунтується на механізмі утворення мРНК. Віруси мають синтезувати мРНК з власних геномів для утворення білків і реплікації своєї нуклеїнової кислоти, однак кожна родина вірусів має власний механізм здійснення цього. Вірусні геноми бувають одноланцюговими (ол) або дволанцюговими (дл), ДНК — або РНК-вмісними, можуть використовувати або не використовувати зворотну транскриптазу. Крім того, одноланцюгові РНК-віруси можуть мати позитивний (+) або негативний (-) ланцюг РНК у складі свого геному.
Ця система містить сім основних груп:
- (I) Віруси, що містять дволанцюгову ДНК і не мають РНК-стадії (наприклад, герпесвіруси, поксвіруси, паповавіруси, мімівірус).
- (II) Віруси, що містять одноланцюгову молекулу ДНК (наприклад, парвовіруси). У цьому разі ДНК завжди позитивної полярності.
- (III) Віруси, що містять дволанцюгову РНК (наприклад, ротавіруси).
- (IV) Віруси, що містять одноланцюгову молекулу РНК позитивної полярності (наприклад, пікорнавіруси, флавівіруси).
- (V) Віруси, що містять одноланцюгову молекулу РНК негативної або подвійної полярності (наприклад, ортоміксовіруси, філовіруси).
- (VI) Віруси, що містять одноланцюгову молекулу РНК позитивної полярності і мають у своєму життєвому циклі стадію синтезу ДНК на матриці РНК, ретровіруси (наприклад, ВІЛ).
- (VII) Віруси, що містять частково дволанцюгову, частково одноланцюгову ДНК і мають у своєму життєвому циклі стадію синтезу ДНК на матриці РНК, ретроїдні віруси (наприклад, вірус гепатиту B).
Подальший поділ здійснюють на основі таких ознак як структура геному (наявність сегментів, кільцева або лінійна молекула), генетична схожість з іншими вірусами, наявність ліпідної оболонки, таксономічна належність організму-хазяїну тощо.
Роль у захворюваннях людини

Прикладами найвідоміших вірусних захворювань людини є застуда (вона може мати й бактеріальну етіологію), грип, вітряна віспа і простий герпес. Багато важких хвороб, наприклад, геморагічна лихоманка Ебола, СНІД, пташиний грип і тяжкий гострий респіраторний синдром спричиняються вірусами. Відносна здатність вірусу спричиняти захворювання характеризується терміном вірулентність. Деякі захворювання досліджують на наявність вірусів серед чинників, наприклад, можливий зв'язок між людським герпесвірусом 6-го типу і нейрологічними захворюваннями, як-от розсіяний склероз і синдром хронічної втоми. Йдуть суперечки щодо того, що борнавірус, якого раніше вважали збудником нейрологічних захворювань у коней, можливо, породжує психічні розлади у людей.
Віруси мають різні механізми, що спричинюють хворобу у хазяїна, і ці механізми сильно залежать від виду. Такий механізм на клітинному рівні включає насамперед лізис клітин, що призводить до їхньої смерті. У багатоклітинних організмів після загибелі значної кількості клітин починає страждати організм у цілому. Хоча віруси підривають нормальний гомеостаз, призводячи до захворювання, вони можуть існувати всередині організму і відносно нешкідливо. Як приклад можна навести здатність вірусу простого герпесу першого типу перебувати в стані спокою всередині тіла людини. Такий стан називають латентністю. Він характерний для вірусів герпесу, у тому числі вірусу Епштейна — Барр, що спричинює інфекційний мононуклеоз, а також вірусу, який спричиняє вітрянку і оперізувальний герпес. Більшість людей перехворіли принаймні одним з цих типів вірусу герпесу. Однак такі латентні віруси можуть бути корисними, оскільки наявність їх може спричинити імунну відповідь проти бактеріальних патогенів, наприклад, чумної палички (Yersinia pestis).
Деякі віруси можуть спричинити довічні або хронічні інфекції, коли вірус і далі розмножується в тілі організму, попри його захисні механізми. Так відбувається, наприклад, з інфекціями, спричиненими вірусами гепатиту B і C. Хронічно хворих людей називають носіями, оскільки вони виступають у ролі резервуара для заразного вірусу. Якщо в популяції є висока частка носіїв, то у цьому випадку кажуть про епідемію.
Епідеміологія
Вірусна епідеміологія є частиною медичної науки, що вивчає передавання та контроль вірусних інфекцій серед людей. Передавання вірусів може здійснюватися вертикально, тобто від матері до дитини, або горизонтально, тобто від людини до людини. Прикладами вертикальної передачі є вірус гепатиту B і ВІЛ, коли малюк народжується вже зараженим. Іншим, рідкіснішим, прикладом є вірус вітряної віспи та оперізувального герпесу, який, хоча й спричинює відносно слабкі інфекції серед дорослих людей, може вбити ембріона і новонародженого малюка.
Горизонтальна передача є найпоширенішим механізмом поширення вірусу в популяції. Передавання може здійснюватися: коли передаються рідини організму під час статевого акту, наприклад, у ВІЛ; через кров під час переливання зараженої крові або користування брудним шприцом, наприклад, у вірусу гепатиту C; передавання Слина губами, наприклад, у вірусу Епштейна — Барр; проковтування зараженої води або їжі, наприклад, у норовірусу; коли вдихається повітря, в якому є віріони, наприклад, вірус грипу; комахами, наприклад, комарами, які ушкоджують шкіру хазяїна, наприклад, гарячка денге. Швидкість передачі вірусної інфекції залежить від кількох факторів, до яких відносять щільність популяції, кількість чутливих людей (тобто таких, що не мають імунітету), якість охорони здоров'я і погоду.
Епідеміологію використовують, аби призупинити поширення інфекції в популяції під час спалаху вірусного захворювання. Роблять контрольні заходи, основані на знанні того, як поширюється вірус. Важливо знайти джерело (або джерела) спалаху та ідентифікувати вірус. Коли вірус визначено, буває можливим зупинити інфекцію за допомогою вакцин. Якщо вакцини недоступні, можуть бути ефективними санація та дезінфекція. Часто заражених людей ізолюють від решти суспільства, тобто вірус поміщають у карантин. Щоб взяти під контроль спалах ящуру у Великій Британії 2001 року, було зарізано тисячі корів. У більшості інфекцій людини і тварин є інкубаційний період, протягом якого не виявляється жодних симптомів інфекції. Інкубаційний період вірусних захворювань може тривати від кількох днів до тижнів. Часто перекривається з ним, але переважно йде після інкубаційного періоду — період передавання інфекції, коли заражена людина чи тварина є заразною й може заразити інших людей або тварин. Цей період також відомий для багатьох інфекцій, і знання тривалості обох періодів є важливим для контролю над спалахами. Якщо спалах призводить до незвичайно високої кількості випадків захворювання в популяції або регіоні, то його називають епідемією. Якщо спалахи поширюються в глобальному масштабі, то кажуть про пандемію.
Епідемії та пандемії
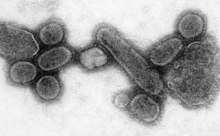
Чисельність корінного населення Америки сильно зменшилась внаслідок заразних захворювань, зокрема, віспи, що їх завезли в Америку європейські колонізатори. За деякими оцінками, іноземні хвороби після прибуття Колумба в Америку, вбили близько 70 % від всього корінного населення. Збитки, завдані цими хворобами аборигенам, допомогли європейцям витіснити і підкорити їх.
Пандемія — це епідемія світового масштабу. Епідемія іспанського грипу 1918 року, що тривала до 1919 року, належить до 5-ї категорії пандемій вірусу грипу. Її спричинив надзвичайно агресивний і смертоносний вірус грипу A. Його жертвами часто ставали здорові дорослі люди, на відміну від більшості спалахів грипу, які вражали переважно дітей і підлітків, людей старшого покоління та інших ослаблених людей. За старими оцінками, іспанський грип забрав 40-50 млн життів, а за сучасними оцінками ця цифра наближається до 100 млн, тобто 5 % тодішнього населення Землі.
Більшість дослідників вважають, що ВІЛ з'явився в Субсахарській Африці у XX столітті. Нині епідемія СНІД має масштаб пандемії. За оцінками, нині 38,6 мільйонів людей на землі заражені ВІЛ. За оцінками Об'єднаної програми Організації Об'єднаних Націй з ВІЛ/СНІД та Всесвітньої організації охорони здоров'я, від СНІДу (останньої стадії ВІЛ-інфекції) померло понад 25 мільйонів осіб з моменту реєстрації першого випадку захворювання 5 червня 1981 року, що робить його однією з найбільш руйнівних епідемій за всю документовану історію. 2007 року зареєстровано 2,7 млн випадків зараження ВІЛ і 2 млн смертей від пов'язаних з ВІЛ захворювань.
Кілька високолетальних вірусних патогенів належать до родини філовірусів. Філовіруси є філаментоподібними вірусами, що спричиняють геморагічну гарячку, до них також відносять збудника геморагічної гарячки Ебола і вірус Марбург. Вірус Марбург привернув широку увагу преси у квітні 2005 року через спалах в Анголі. Цей спалах тривав з жовтня 2004 року і аж до 2005 року й увійшов в історію як найжахливіша епідемія будь-якої геморагічної гарячки.

Злоякісні пухлини
Віруси можуть спричиняти злоякісні новоутворення (зокрема, рак печінки або саркому Капоші) у людини та інших видів, хоча вони виникають лише у незначної частини інфікованих. Пухлинородні віруси належать до різних родин; серед них і РНК-, і ДНК-вмісні віруси, тому єдиного типу «онковірус» не існує (застарілий термін, спочатку застосовуваний до ретровірусів, що швидко трансформуються). Розвиток раку визначається багатьма факторами, як-от імунітет хазяїна і його мутації. До вірусів, що здатні породити рак у людини, відносять деяких представників папіломавірусу людини, вірусу гепатиту B і C, вірусу Епштейна — Барр, герпесвірусу саркоми Капоші і людського T-лімфотропного вірусу. Зовсім нещодавно відкритим вірусом раку людини є поліомавірус клітин Меркеля, який у більшості випадків спричинює рідкісну форму раку шкіри, званого карциномою клітин Меркеля. Віруси гепатиту можуть спричинити хронічну вірусну інфекцію, яка призводить до раку печінки. Зараження людським T-лімфотрофним вірусом може призвести до тропічного спастичного параперезу і зрілого лейкозу Т-клітин. Людські папіломавіруси можуть спричиняти рак шийки матки, шкіри, ануса і статевого члена. Серед герпесвірусів герпесвірус саркоми Капоші спричиняє саркому Капоші і лімфому порожнини тіла, вірус Епштейна — Барр — лімфому Беркітта, лімфогранулематоз, порушення B-лімфопроліферації та назофарингеальну карциному. Поліомавірус клітин Меркеля близький до вірусу SV40 і поліомавірусів мишей, яких понад 50 років використовували як тваринних моделей для вивчення вірусного раку.
Захисна реакція хазяїна
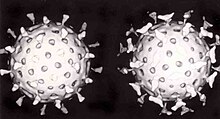
Першою захисною лінією організму проти вірусу є вроджений імунітет. До нього належать клітини та інші механізми, що забезпечують неспецифічний захист. Це означає, що клітини вродженого імунітету розпізнають і реагують на патогени загальними способами, однаково щодо всіх патогенів, але, на відміну від набутого імунітету, вроджений імунітет не дає хазяїну тривалого й надійного захисту.
Важливим природним способом захисту організму еукаріотів проти вірусів є РНК-інтерференція. Стратегія реплікації багатьох вірусів передбачає наявність стадії дволанцюгової РНК. Для боротьби з подібними вірусами, клітина має систему неспецифічної деградації одно — і дволанцюгових РНК. Коли подібний вірус проходить у клітину і вивільняє геномну РНК в цитоплазму, білковий комплекс дайсер пов'язує і розриває вірусну РНК на короткі фрагменти. Задіюється біохімічний шлях, званий RISC, який руйнує вірусну РНК, і перешкоджає розмноженню вірусу. Ротавіруси здатні уникнути РНК-інтерференції, зберігаючи частину капсиду навіть всередині клітини і випускаючи новостворені мРНК через шпарини у внутрішньому капсиді. Геномна дволанцюгова РНК ж залишається всередині нього.
Коли система набутого імунітету у хребетних стикається з вірусом, вона утворює специфічні антитіла, що приєднуються до вірусу і часто роблять його безпечним. Це називається гуморальним імунітетом. Найважливішими є два типи антитіл. Перший, званий IgM, дуже ефективно нейтралізує віруси, але його утворюють клітини імунної системи лише протягом кількох тижнів. Синтез другого — IgG — триває невизначено довго. Присутність IgM у крові господаря свідчить про наявність гострої інфекції, тоді як IgG свідчить про інфекцію, перенесену в минулому. Саме кількість IgG вимірюють коли тестують на імунітет. Антитіла можуть залишатися ефективним захисним механізмом навіть тоді, коли вірусу вдається пройти в клітину. Клітинний білок TRIM21 може прикріплювати антитіла до поверхні вірусних частинок. Це спричинює подальше руйнування вірусної частинки ферментами клітинної протеасомної системи.
Другий захисний механізм хребетних проти вірусів називається клітинним імунітетом і включає імунні клітини, відомі як T-лімфоцити. Клітини тіла постійно несуть короткі фрагменти власних білків на своїх поверхнях, і, якщо Т-лімфоцити розпізнають тут підозрілі вірусні фрагменти, клітина-хазяїн руйнується клітинами, що називаються Т-кілерами, і починається утворення специфічних до вірусу T-лімфоцитів. Такі клітини, як макрофаги, спеціалізуються на презентації антигенів.
Важливою захисною реакцією хазяїна є вироблення інтерферону. Інтерферон — це білок. Його роль в імунітеті — комплексна, зрештою він зупиняє вірус, припиняючи утворення нових вірусів ураженими клітинами, вбиваючи їх та їхніх близьких сусідів. З убитих вірусом клітин у навколишнє середовище попадає деяка кількість фрагментів двониткової РНК. На поверхні клітин існують білкові рецептори, які впізнають такий РНК і передають сигнал про таку взаємодію у ядро, де депресується структурний ген інтерферону[джерело?].
Не проти всіх вірусів утворюється така захисна імунна відповідь. ВІЛ вдається уникнути імунної відповіді, постійно змінюючи послідовність амінокислот поверхневих білків віріона. Такі стійкі віруси проскочують повз імунну систему, ізолюючись від імунних клітин, блокуючи презентацію антигенів, завдяки стійкості до цитокінів, ухиляючись від природних кілерів, зупиняючи апоптоз клітин-господарів, а також за рахунок антигенної мінливості. Інші віруси, звані нейротропними вірусами, поширюються серед нервових клітин, тобто там, де імунна система не в змозі дістатися до них[джерело?].
Профілактика й лікування
Оскільки віруси використовують для розмноження природні метаболічні шляхи клітин-господарів, то їх складно знищити без застосування препаратів, токсичних для самих клітин-господарів. Найефективнішими медичними заходами проти вірусних інфекцій є вакцинації, що створюють імунітет до інфекції, і противірусні препарати, що вибірково інгібують реплікацію вірусів.
Вакцини
Вакцинація є дешевим і ефективним способом запобігання вірусних інфекцій. Вакцини для запобігання вірусних інфекцій застосовували ще задовго до відкриття самих вірусів. Їх застосування допомогло різко знизити протікання та смертністю від вірусних інфекцій, як-от поліомієліт, кір, свинка і краснуха. З допомогою вакцинації віспу викорінено. За допомогою вакцин можна запобігти понад 30 вірусних інфекцій у людини, а ще більше вакцин використовують для запобігання вірусних захворювань тварин. Вакцини можуть містити ослаблені й убиті віруси, а також вірусні білки (антигени). Живі вакцини містять ослаблені форми вірусів, які не спричинюють хвороби, але породжують імунну відповідь. Такі віруси називають атенуйованими. Живі вакцини можуть становити небезпеку для людей зі слабким імунітетом (тобто мають імунодефіцит), бо навіть ослаблений вірус у них може спричинити вихідне захворювання. Для виробництва т. зв. субодиничних вакцин використовують біотехнології та генетичну інженерію. У таких вакцинах використовують тільки капсидні білки вірусів. Прикладом таких вакцин є вакцина проти вірусу гепатиту B. Субодиничні вакцини нешкідливі для людей з імунодефіцитом, тому що вони не можуть спричинити захворювання. Вакцина проти жовтої гарячки, що містить ослаблений штам 17D, мабуть, є найбільш ефективною та безпечною з будь-коли створених вакцин.
Противірусні препарати


Противірусні препарати часто є аналогами нуклеозидів. Вони вбудовуються в геном вірусу під час реплікації, і на цьому життєвий цикл вірусу зупиняється, оскільки новосинтезована ДНК недіяльна. Це викликано тим, що в аналогів відсутні гідроксильні групи, які разом з атомами фосфору з'єднуються і формують жорсткий «кістяк» молекули ДНК. Це називається ланцюговою термінацією ДНК. Приклади аналогів нуклеозидів — ацикловір, що застосовується проти інфекцій, які спричинюють вірус простого герпесу, і ламівудин (проти ВІЛ та вірусу гепатиту B). Ацикловір — один з найстаріших і найчастіше призначуваних противірусних препаратів.
Інші застосовувані противірусні препарати мають мішенню різні стадії життєвого циклу вірусів. ВІЛ, щоб стати повністю заразним, потребує протеолітичного ферменту, відомого як ВІЛ-1 протеаза. На підставі цього розроблено великий клас препаратів, званих інгібіторами протеази, що інактивують цей фермент.
Гепатит C спричиняється РНК-вмісним вірусом. У 80 % інфікованих людей інфекція має хронічний характер, і без лікування вони залишаться зараженими до кінця своїх днів. Однак на початку XXI століття застосовують ефективні ліки, що складаються з нуклеозидного аналога рибавірину, комбінованого з інтерфероном. Для лікування хронічних носіїв гепатиту B розроблено схоже лікування з використанням ламівудину.
Вірусні захворювання у різних організмів
Віруси вражають усе клітинне життя, але, попри повсюдне поширення вірусів, кожен вид клітинних організмів має свою низку вірусів, що вражають, часто тільки цей вид. Деякі віруси, звані сателітами, можуть розмножуватися лише в клітинах, вже заражених іншим вірусом.
Віруси тварин
У тварин вірусні інфекції породжують імунну відповідь, яка найчастіше призводить до знищення хвороботворного вірусу. Імунну відповідь також можна спричинити вакцинами, що дають діяльний набутий імунітет проти саме цієї вірусної інфекції. Однак деякі віруси, зокрема ВІЛ і збудники вірусних гепатитів, здатні проскочити повз імунну відповідь, породжуючи хронічну хворобу. Антибіотики не діють на віруси, проте було розроблено декілька противірусних препаратів (див. вище).
Віруси є важливими патогенами домашньої худоби. Вони спричиняють, наприклад, ящур і «синій язик». Домашні тварини, приміром, коти, собаки і коні, якщо їх не вакцинувати, є чутливими до серйозних вірусних хвороб. Собачий парвовірус — це маленький ДНК-вмісний вірус, що часто вбиває цуценят. Однак більшість вірусів нешкідливо співіснують зі своїми хазяями, без жодних ознак або симптомів хвороби.
Віруси безхребетних
На частку безхребетних припадає близько 80 % всіх відомих видів тварин, тому немає нічого дивного в тому, що вони приховують у собі величезну кількість вірусів різних типів. Найбільш вивчені віруси, що вражають комах, але навіть тут доступна про них інформація має фрагментарний характер. Втім, останнім часом описано вірусні захворювання і в інших безхребетних. Ці віруси й далі маловивчені, і деякі повідомлення про відкриття слід приймати обережно, поки вірусна природа цих хвороб не буде остаточно доведена. Крім того, необхідно також перевірити інфективність ізольованих вірусів щодо неінфікованих господарів того ж виду, у якого ці віруси виявлено.
Нині виділено окрема родина вірусів, що вражають головним чином членистоногих, особливо комах, які живуть у водних і вологих середовищах: іридовіруси, від англ. Invertebrate iridescent viruses — «веселкові віруси безхребетних»; такого кольору зразки уражених комах). Це ікосаедричні частинки діаметром 120—180 нм, що містять внутрішню ліпідну мембрану і геном у вигляді дволанцюгової ДНК, що містить 130—210 тис. пар нуклеотидів.
Інші віруси, що вражають комах: родина Baculoviridae, підродина Entomopoxvirinae родина Поксвіруси, рід Densovirus родина Parvoviridae, деякі віруси родини Рабдовіруси, Реовіруси, Пікорнавірус.
Як і всі безхребетні, медоносна бджола чутлива до багатьох вірусних інфекцій.
Віруси рослин

Існує багато типів вірусів рослин. Часто вони спричиняють зниження врожайності, завдаючи значних збитків сільському господарству, тому контроль таких вірусів є дуже важливим з економічного погляду. Віруси рослин часто поширюються від рослини до рослини організмами, відомими як переносники. Зазвичай це комахи, але ними можуть бути також гриби, черви-нематоди і одноклітинні організми. Якщо контроль вірусу рослин визнають економічно вигідним, наприклад, у випадку багаторічних фруктових дерев, зусилля спрямовують на усунення переносників або альтернативних господарів, наприклад, бур'янів. Віруси рослин не можуть вражати людину та інших тварин, бо вони можуть розмножуватися лише у живих рослинних клітинах.
Рослини мають складні та ефективні механізми захисту від вірусів. Найефективнішим механізмом є наявність так званого гена стійкості (R від англ. resistance — «стійкість»). Кожен R-ген відповідає за стійкість до окремого вірусу і спричинює загибель клітин, сусідніх з ураженої, що неозброєним оком видно як велика пляма. Це зупиняє розвиток хвороби внаслідок зупинки поширення вірусу. Іншим ефективним методом є РНК-інтерференція. Уражені вірусом рослини часто починають виробляти природні противірусні речовини, як-от саліцилова кислота, оксид азоту та активні форми кисню.
Віруси рослин і створені на їх основі вірусоподібні частинки (VLPs) знайшли застосування в біотехнології та нанотехнології. Капсиди більшості вірусів рослин мають просту і стійку структуру, і вірусні частинки можуть вироблятися у величезних кількостях як ураженою рослиною, так і різними гетерологічними системами. Віруси рослин можуть змінюватися хімічно і генетично, оточуючи оболонкою чужорідні частинки, а також здатні вбудовуватися в надмолекулярні структури, що вможливлює їх застосування в біотехнологіях.
Віруси грибів
Віруси грибів називають міковірусами. Нині віруси виділено в 73 видів з 57 родів, що належать до 5 класів, але, імовірно, у нешкідливому стані віруси існують у більшості грибів. Загалом ці віруси мають вигляд круглих частинок діаметром 30-45 нм, що складаються з багатьох субодиниць єдиного білка, складених навколо геному представленого дволанцюговою РНК. Зазвичай, віруси грибів відносно нешкідливі. Деякі грибні штами можуть вражатися багатьма вірусами, але більшість міковірусів тісно пов'язані зі своїм єдиним хазяїном, від якого передаються його нащадкам. Над класифікацією вірусів грибів нині працює спеціально створений відділ у складі ICTV. Нині він визнає 3 родини вірусів грибів, а найбільш вивчені міковіруси належать до родини Totiviridae
Встановлено, що антивірусна активність пеніцилінових грибів зумовлена індукцією інтерферону дволанцюгової РНК від вірусів, що вражають гриби.
Якщо ж вірус, потрапляючи в гриб, проявляє свою вірулентність, то реакція гриба на це може бути різною: зниження або підвищення вірулентності у патогенних видів, дегенерація грибниці та плодових тіл, зміна забарвлення, придушення спороношення. Некапсидовані вірусні РНК передаються через анастомози незалежно від мітохондрій.
Вірусні захворювання можуть завдавати шкоди грибівничим підприємствам, наприклад, спричинити порудіння плодових тіл печериці, зміну забарвлення у зимового опенька, що знижує їх комерційну цінність. Віруси, що породжують гіповірулентність грибів-патогенів, можна використовувати для боротьби із захворюваннями рослин.
Віруси протистів
До вірусів протистів відносять віруси, що вражають одноклітинних еукаріотів, не включених у царство тварини, рослини або гриби. Деякі з відомих нині вірусів протистів:
| Назва вірусу (рід) | Систематичне положення (родина) |
Уражений протист |
|---|---|---|
| Dinornavirus | Alvernaviridae | Heterocapsa circularisquama |
| Endornavirus | Endornaviridae | Фітофтора |
| Labyrnavirus | Labyrnaviridae | Aurantiochytrium |
| Marnavirus | Marnaviridae | Heterosigma akashiwo |
| Marseillevirus | Marseilleviridae | Амеба |
| Мімівірус | Mimiviridae | Acanthamoeba polyphaga |
| Chlorovirus | Phycodnaviridae | Paramecium bursaria |
| Coccolithovirus | Phycodnaviridae | Emiliania huxleyi |
| Prasinovirus | Phycodnaviridae | Micromonas pusilla |
| Prymnesiovirus | Phycodnaviridae | Chrysochromulina brevifilum |
| Raphidovirus | Phycodnaviridae | Heterosigma akashiwo |
| Cryspovirus | Partitiviridae | Cryptosporidium parvum |
| Hemivirus | Pseudoviridae | Volvox carteri |
| Pseudovirus | Pseudoviridae | Physarum polycephalum |
| Mimoreovirus | Реовіруси | Micromonas pusilla |
| Giardiavirus | Totiviridae | Giardia lamblia |
| Leishmaniavirus | Totiviridae | Лейшманія |
| Trichomonasvirus | Totiviridae | Трихомонада вагінальна |
| Bacilladnavirus | Не визначено |
Chaetoceros salsugineum Rhizosolenia setigera |
| Dinodnavirus | Не визначено | Heterocapsa circularisquama |
| Rhizidiovirus | Не визначено | Rhizidiomyces |
Багато вірусів найпростіших мають незвичайно великі розміри. Наприклад, геном Marseillevirus, вперше виділений з амеби, має геном розміром 368 КБ, а Mamavirus, що вражає протиста Acanthamoeba, за розміром перевершує навіть мімівірус (а його капсид сягає близько 500 нм в діаметрі) та деякі бактерії. Також до гігантських вірусів належить вірус, що вражає поширеного морського протиста Cafeteria roenbergensis (англ. Cafeteria roenbergensis virus, CroV).
Віруси бактерій
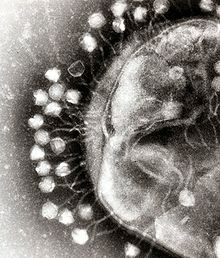
Бактеріофаги є поширеною й різноманітною групою вірусів, що чисельніша у водних середовищах проживання — в океанах цих вірусів у понад 10 разів більше, ніж бактерій, досягаючи чисельності в 250 млн вірусів на мілілітр морської води. Ці віруси вражають специфічні для кожної групи бактерії, зв'язуючись з клітинними рецепторами на поверхні клітини і потім проходячи всередину неї. Протягом короткого проміжку часу (іноді лічених хвилин) бактеріальна полімераза починає транслювати вірусну мРНК в білки. Ці білки або входять до складу віріонів, що накопичуються всередині клітини, або є допоміжними білками, що допомагають складанню нових віріонів, або спричинюють лізис клітини. Вірусні ферменти спричинюють руйнування клітинної мембрани, і, як у випадку фага Т4, лише через 20 хвилин після проникнення в клітину виникають понад триста бактеріофагів.
Головним механізмом захисту бактеріальних клітин від бактеріофагів є утворення ферментів, що руйнують чужорідну ДНК. Ці ферменти, звані ендонуклеазами рестрикції, «розрізають» вірусну ДНК, впорснуту всередину клітини. Бактерії також використовують систему, звану CRISPR, яка зберігає інформацію про геноми вірусів, з якими бактерія стикалася раніше, і це дозволяє клітині блокувати реплікацію вірусу за допомогою інтерференції РНК. Ця система забезпечує набутий імунітет бактеріальної клітини.
Бактеріофаги можуть виконувати і корисну для бактерій функцію, приміром, саме бактеріофаг, що заражає дифтерійні палички, кодує ген їхнього токсину, потрібного цим бактеріям і такого небезпечного для людини.
Віруси архей
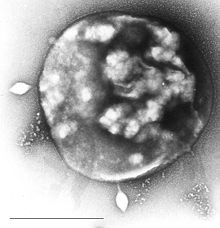
Деякі віруси розмножуються всередині архей: це дволанцюгові ДНК-вмісні віруси незвичайної, часом унікальної форми. Найбільш детально вони вивчені у термофільних архей, зокрема, порядків Sulfolobales і Thermoproteales. Заходами захисту проти цих вірусів можуть бути РНК-інтерференція від повторюваних послідовностей ДНК в геномах архей, споріднених генам вірусів.
Віруси вірусів

Під час вивчення вірусних фабрик мімівірусу виявлено, що на них збираються невеликі віріони й іншого вірусу, якого назвали Супутником. Супутник, найімовірніше, сам не здатен заражати клітини амеб (які є хазяями мімівірусу) і розмножуватися в них, але може робити це спільно з мама — або мімівірусом, що класифікує його як вірус-сателіт. Супутник став першим відомим вірусом-сателітом, що містить дволанцюгову ДНК і розмножується в еукаріотичних клітинах. Однак автори роботи пропонують розглядати його не просто як сателіт, а як вірофаг (вірус вірусу) за аналогією з бактеріофагами (вірусами бактерій). Реплікація і вірусів-сателітів, і вірофагів залежить від іншого вірусу й клітини-хазяїна. Однак для реплікативного циклу вірофагів характерні три унікальні особливості. 1) Відсутня ядерна фаза реплікації. 2) Реплікація вірофагів відбувається у вірусних фабриках гігантських ДНК-вмісних вірусів-господарів. 3) Вірофаги залежать від ферментів, синтезованих вірусами-господарями, але не клітинами-господарями. Таким чином, вірофагів вважають паразитами гігантських ДНК-вмісних вірусів, наприклад, мімівірусів і фікоднавірусів. При цьому синтез капсидних білків вірофагів (як і синтез білків усіх відомих вірусів) повністю залежить від трансляційного апарату клітини-хазяїна. Хоча суворого доказу ще немає, деякі факти свідчать на користь того, що Супутник справді є вірофагом. Наприклад, у його геномі наявні регуляторні елементи, характерні для мімівірусу і впізнанні його транскрипційним апаратом (послідовності, близькі до пізнього промотора мімівірусу, сигнали поліаденілування). Крім того, наявність Супутника знижує продуктивність розмноження мімівірусу: лізис клітини-хазяїна відбувається із затримкою, і утворюються дефектні віріони мімівірусу. За даними на 2016 рік, з культивованих клітин було ізольовано п'ять вірофагів. Ще 18 вірофагів описано на основі даних метагеномного аналізу (геноми двох з них майже повністю секвеновано).
Роль вірусів у біосфері
Віруси є найпоширенішою за чисельністю формою існування органічної матерії на планеті. Вони відіграють важливу роль у регуляції чисельності популяцій деяких видів живих організмів (наприклад, вірус дикування з періодом у декілька років скорочує чисельність песців у кілька разів).
Іноді віруси утворюють з тваринами симбіоз. Так, наприклад, отрута деяких паразитичних ос містить структури, звані полі-ДНК-вірусами (Polydnavirus, PDV), які мають вірусне походження.
Однак основна роль вірусів у біосфері пов'язана з їх діяльністю у водах океанів і морів.
Роль у водних екосистемах
Віруси — це найпоширеніша форма життя в океані, їх концентрація сягає 10 млн вірусів на 1 мм2 поверхні моря. Чайна ложка морської води містить близько мільйона вірусів. Вони необхідні для регуляції прісноводних і морських екосистем. Значна частина цих вірусів є бактеріофагами, нешкідливими для рослин і тварин. Вони вражають і руйнують бактерії у водному мікробному співтоваристві, таким чином, беручи участь у важливому процесі кругообігу вуглецю в морському середовищі. Органічні молекули, які звільнилися з бактеріальних клітин завдяки вірусам, стимулюють ріст нових бактерій і водоростей.
Мікроорганізми становлять понад 90 % біомаси в морі. За оцінками, кожен день віруси вбивають близько 20 % цієї біомаси, а кількість вірусів в океанах у 15 разів перевищує кількість бактерій і архей. Віруси є головними агентами, що спричиняють швидке припинення цвітіння води, яке вбиває інше життя в морі, завдяки загибелі водоростей, що спричинюють його. Чисельність вірусів зменшується з віддаленням від берега і зі збільшенням глибини, оскільки там менше організмів-хазяїв.
Значення морських вірусів дуже велике. Регулюючи процес фотосинтезу, вони відіграють другорядну роль у скороченні кількості вуглекислого газу в атмосфері приблизно на 3 гігатонни вуглецю за рік.
Як і інші організми, морські ссавці сприйнятливі до вірусних інфекцій. У 1988 і 2002 роках тисячі звичайних тюленів були вбиті параміксовірусом Phocine distemper virus. У популяціях морських ссавців циркулює безліч інших вірусів, у тому числі каліцивіруси, герпесвіруси, аденовіруси та парвовіруси.
Роль в еволюції
Віруси є важливим природним засобом перенесення генів між різними видами, що породжує генетичне різноманіття і направляє еволюцію. Вважають, що віруси відіграли центральну роль у ранній еволюції, ще до розбіжності бактерій, архей і еукаріотів, у часи останнього універсального загального предка життя на Землі. Віруси й донині є одним з найбільших живих сховищ недослідженого генетичного різноманіття на Землі.
Застосування
У науках про життя та медицині

Віруси мають важливе значення для досліджень молекулярної і клітинної біології, бо вони є простими системами, які можна використовувати для управління і вивчення функціонування клітин. Вивчення і використання вірусів дало цінну інформацію про різні аспекти клітинної біології. Наприклад, віруси застосовували в генетичних дослідженнях, і вони допомогли дійти розуміння ключових механізмів молекулярної генетики, таких як реплікація ДНК, транскрипція, процесинг РНК, трансляція, транспорт білків.
Генетики часто використовують віруси як вектори для введення генів у досліджувані клітини. Це дозволяє змусити клітину виробляти чужі речовини, а також вивчити ефект від введення нового гена в геном. Аналогічно у віротерапії віруси використовують як вектори для лікування різних хвороб, тому що вони вибірково діють на клітини і ДНК. Це дає надії, що віруси зможуть допомогти в боротьбі з раком і знайдуть своє застосування у генотерапії. Деякий час східноєвропейські вчені застосовували фагової терапії як альтернативу антибіотиків, і інтерес до таких методів зростає, оскільки нині у деяких патогенних бактерій виявлено високу стійкість до антибіотиків.
Біосинтез зараженими клітинами чужорідних білків лежить в основі деяких сучасних промислових способів отримання білків, наприклад, антигенів. Нещодавно промисловим способом отримано деякі вірусні вектори і лікарські білки, нині вони проходять клінічні та доклінічні випробування.
У матеріалознавстві і нанотехнологіях
Сучасні напрямки в нанотехнології обіцяють значно урізноманітнити застосування вірусів. З погляду матеріалознавців, віруси можна розглядати як органічні наночастинки. Їхня поверхня несе спеціальні пристосування для подолання біологічних бар'єрів клітини-хазяїна. Точно визначено форму і розмір вірусів, а також кількість і природу функціональних груп на поверхні. По суті, віруси часто використовують в матеріалознавстві як «риштування» для ковалентно пов'язаних поверхневих модифікацій. Одна з відмінних якостей вірусів — те, що вони спеціально «підігнані» спрямованою еволюцією під клітини, що є хазяями. Потужні методи, розроблені біологами, лягли в основу інженерних прийомів у наноматеріалах, відкривши тим самим широку сферу застосування вірусів, що виходить далеко за межі біології та медицини.
Завдяки їхнім розмірам, формі й добре вивченій хімічній структурі віруси використовували як шаблони для організації матеріалів на нанорівні. Прикладом такої нещодавньої роботи можуть бути дослідження, проведені Дослідницькою лабораторією Навалю у Вашингтоні (округ Колумбія) з використанням вірусу мозаїки коров'ячого гороху (англ. Cowpea Mosaic Virus (CPMV)) для посилення сигналів у сенсорах з ДНК-мікрочіпами. В цьому випадку вірусні частинки поділяли частинки флуоресцентних барвників, які використовувалися для передачі сигналу, запобігаючи, таким чином, скупченню нефлуоресцентних димерів, які виступають як гасителі сигналу. Іншим прикладом використання CPMV є застосування його як нанорозмірного зразка для молекулярної електроніки.
Штучні віруси
Багато вірусів можна отримати de novo, тобто з нуля, а перший штучний вірус отримали 2002 року. Попри деякі неправильні трактування, при цьому синтезується не сам вірус, а його геномна ДНК (у разі ДНК-вмісних вірусів) або комплементарна копія ДНК геному (у разі РНК-вірусів). У вірусів багатьох родин штучна ДНК або РНК (остання виходить шляхом зворотної транскрипції синтетичної комплементарної ДНК), якщо її ввести в клітину, проявляє інфекційні властивості. Іншими словами, вони містять всю необхідну інформацію для утворення нових вірусів. Цю технологію нині використовують для розробки вакцин нового типу. Можливість створювати штучні віруси має далекосяжні наслідки, оскільки вірус не може вимерти, поки відома його геномна послідовність і є чутливі до нього клітини. В наші дні повні геномні послідовності 2408 різних вірусів (в тому числі віспи) є в публічному доступі в онлайн-базі даних, підтримуваній Національними інститутами охорони здоров'я США.
Віруси як зброя
Спроможність вірусів спричиняти спустошливі епідемії серед людей породжує тривогу, що віруси можна застосувати як біологічну зброю. Додаткові побоювання викликало успішне відтворення вірусу іспанського грипу в лабораторії. Іншим прикладом може бути вірус віспи. Упродовж усієї історії він спустошував безліч країн аж до його остаточного викорінення. Офіційно зразки вірусу віспи зберігаються лише в двох місцях у світі — у двох лабораторіях в Росії і США. Побоювання, що його можуть використати як зброю, не зовсім безпідставні; вакцина проти віспи іноді має важкі побічні ефекти — в останні роки до офіційно оголошеного викорінення вірусу більше людей серйозно захворіли через вакцини, ніж від вірусу, тому вакцинацію проти віспи більше не практикують повсюдно. Через це більша частина сучасного населення Землі практично не має стійкості до віспи.
У масовій культурі
У фільмах та інших творах світ інфекційних захворювань, зокрема вірусних, рідко показують достовірно. Крім фільмів-біографій вчених і фільмів, що розповідають про великі епідемії минулого, у більшості з них центральною подією є спалах невідомого хвороботворного агента, поява якого стала наслідком акту біотероризму, інциденту в лабораторії, або ж він потрапив з космосу.
У літературі
Вірусна інфекція закладена в основу таких творів (список неповний):
- Кодзі Судзукі. «Дзвінок».
- Кир Буличов. «Лілова куля».
- Стівен Кінг. «Протистояння».
- Майкл Крайтон. «Штам «Андромеда»».
- Річард Метісон. «Я — легенда».
- Джек Лондон. «Шарлатова чума».
- Ден Браун. «Інферно».
У кінематографі
Спалах незвичайної вірусної інфекції лежить в основі сюжету таких художніх фільмів і серіалів:
- «Годинник Пандори» (1996)
- «28 днів потому» (2003)
- «28 тижнів потому» (2007)
- Штам «Андромеда». Цей фільм, знятий за однойменною повістю Майкла Крайтона, можна назвати найбільш точним у науковому сенсі.
- «12 мавп» (1995)
- «Оселя зла» (2002) і його продовження.
- «Епідемія» (1995)
- «Лілова куля» (1987)
- «Носії» (2009)
- «Я — Легенда» (2007)
- «Зараза» (2011)
- «Карантин» (2008)
- «Карантин 2: Термінал» (2011)
- «Регенезис» (серіал, 2004—2008)
- «Вижити» (серіал 2013)
- «Спіраль» (серіал, 2014—2015)
- «Штам» (серіал, 2014—2015)
- «Останній корабель» (серіал, 2014—2015)
- «Закрита школа» (серіал, 2011—2012)
- «Війна світів Z» (2013)
- «Інферно» (2016)
- «Епідемія (телесеріал)» (2019)
Останніми роками[коли?] віруси нерідко стають «героями» мультфільмів та мультсеріалів, серед яких слід назвати, наприклад, «Осмосис Джонс» (США, 2001), «Оззі і Дрикс» (США, 2002—2004) і «Вірус атакує» (Італія, 2011).
Деякі поширені вірусні захворювання
Див. також
- Інтерференція вірусів
- Вірусологічна теорія еволюції
- Антивірусні препарати
- Прокаріоти
- Бактеріофаг
- Віроїд
- Новий вірус
- Комп'ютерний вірус
- Мобільний вірус
- Медіавірус
Література
- Авторський колектив, за ред. В.П. Широбокова. Медична мікробіологія, вірусологія та імунологія. — Вінниця : Нова Книга, 2011. — 952 с. — 2000 прим. — ISBN 978-966-382-325-6.
- Гудзь С. П., Перетятко Т. Б., Павлова Ю. О. Загальна вірусологія. Навчальний посібник. — Львів: Видавничий центр ЛНУ ім. Івана Франка, 2010. — 264 с. ISBN 9789666137534
- Ташута С. Г. Загальна вірусологія: Посібник. — Київ:, 2004. — 458 с.
- Вірусосфера: від застуди до COVID — навіщо людству віруси / Ф. Раян ; [пер. з англ.: Н. Палій, Р. Свято]. — Київ: Yakaboo Publishing, 2020. — 304 с. ISBN 978-617-7544-70-7
- Fields Virology: Emerging Viruses. Howley, Peter M.; Knipe, David M.; Whelan, Sean/ Lippincott Williams & Wilkins (LWW), 2020.- 1256 p. ISBN 978-1-97-511254-7
- Mayo M. A., Pringle C. R. Virus taxonomy — 1997 // Journal of General Virology. — Microbiology Society, 1998. — No. 79. — P. 649—657. Архівовано з джерела 29 вересня 2007.
- Collier, Leslie; Balows, Albert; Sussman, Max. Topley and Wilson’s Microbiology and Microbial Infections / Mahy, Brian and Collier, Leslie. Arnold. — ninth edition. — Virology, 1998. — Т. 1. — ISBN 0-340-66316-2.
- Dimmock, N.J.; Easton, Andrew J.; Leppard, Keith. Introduction to Modern Virology. — sixth edition. — Blackwell Publishing, 2007. — ISBN 1-4051-3645-6.
- Knipe, David M.; Howley, Peter M.; Griffin, Diane E.; Lamb, Robert A.; Martin, Malcolm A.; Roizman, Bernard; Straus Stephen E. Fields Virology. — Lippincott Williams & Wilkins, 2007. — ISBN 0-7817-6060-7.
- Shors, Teri. Understanding Viruses. — Jones and Bartlett Publishers, 2008. — ISBN 0-7637-2932-9.
- Букринская А. Г. Вирусология. — Медицина, 1986. — 336 с.
Посилання
- ВІРУСИ [Архівовано 10 березня 2016 у Wayback Machine.]
|
| |||||||||||||||||||||||||||
